
B-Cell Receptor Signaling and Beyond: The Role of Igα (CD79a)/Igβ (CD79b) in Normal and Malignant B Cells

1
BIOCEV, First Faculty of Medicine, Charles University, Prumyslova 595, 252 50 Vestec, Czech Republic
2
First Department of Internal Medicine–Hematology, General University Hospital and First Faculty of Medicine, Charles University, 128 08 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2024, 25(1), 10; https://doi.org/10.3390/ijms25010010
Submission received: 13 November 2023
/
Revised: 11 December 2023
/
Accepted: 11 December 2023
/
Published: 19 December 2023
(This article belongs to the Special Issue New Advances in B-cell Lymphoma Biology)
Abstract
:B-cell receptor (BCR) is a B cell hallmark surface complex regulating multiple cellular processes in normal as well as malignant B cells. Igα (CD79a)/Igβ (CD79b) are essential components of BCR that are indispensable for its functionality, signal initiation, and signal transduction. CD79a/CD79b-mediated BCR signaling is required for the survival of normal as well as malignant B cells via a wide signaling network. Recent studies identified the great complexity of this signaling network and revealed the emerging role of CD79a/CD79b in signal integration. In this review, we have focused on functional features of CD79a/CD79b, summarized signaling consequences of CD79a/CD79b post-translational modifications, and highlighted specifics of CD79a/CD79b interactions within BCR and related signaling cascades. We have reviewed the complex role of CD79a/CD79b in multiple aspects of normal B cell biology and how is the normal BCR signaling affected by lymphoid neoplasms associated CD79A/CD79B mutations. We have also summarized important unresolved questions and highlighted issues that remain to be explored for better understanding of CD79a/CD79b-mediated signal transduction and the eventual identification of additional therapeutically targetable BCR signaling vulnerabilities.
1. Introduction
B-cell receptor (BCR) signaling plays a critical role at multiple stages of B cell life cycle [1]. BCR is a hallmark molecule of B cells that determines cellular fate and regulates B cell survival, fitness, activation, development, and transformation into immunoglobulin-secreting cells [2]. Structurally, BCR is composed of a membrane-bound immunoglobulin (Ig) molecule noncovalently linked with Igα (CD79a) and Igβ (CD79b) transmembrane signaling subunits [2,3,4]. BCR has a central role within the adaptive immune system and antigen recognition. Antigen binding triggers antigen induced BCR signaling, activating an array of signaling cascades and cellular processes. These include BCR-mediated antigen capture, recognition, uptake, and processing (also making B cells effective antigen-presenting cells) [5]. Importantly, BCR is also a source of baseline antigen-independent signaling, the so-called “tonic” BCR signaling. Tonic BCR signaling is necessary for correct B-cell development and for survival of normal mature B cells [6]. At the same time, altered BCR signaling has been implicated in the pathogenesis of autoimmune inflammatory disorders and B-cell derived malignancies [7,8,9,10].
In recent years, multiple studies have substantially improved our understanding of molecular mechanisms governing BCR signaling and highlighted the critical role of BCR signaling integrative components CD79a and CD79b. In this review, we focus specifically on functional features of the CD79a/CD79b heterodimeric unit in normal and malignant B cells, as well as on its role in BCR signal regulation. Furthermore, we summarize types and consequences of CD79a/CD79b heterodimer covalent posttranslational modifications, describe the CD79a/CD79b interaction network, and highlight CD79a- and CD79b-associated disruption of BCR signaling in malignant B cells. Reflecting the aim of our review, we focus specifically on signaling events related to CD79a and CD79b molecules, providing only a brief overview of BCR signaling in general.
2. BCR Signaling in Normal B Cells
BCR signaling is of critical importance for normal B cell performance at all different stages of their development. BCR can induce a wide array of cellular responses related to the complexity of the intracellular BCR signaling network. Individual features of BCR signaling are highly dependent on a specific stage of B cell development and activation status [11]. Importantly, pre-BCR, which is structurally similar to BCR but contains a surrogate light chain (SLC) made up of the invariant proteins λ5 (CD179b) and VpreB (CD179a) instead of the Ig light chain in BCR [12], is transiently expressed in early developmental stages and is necessary for pro-B to pre-B transition and pre-B cell expansion [13,14]. Correct assembly and proper SLC replacement with the conventional light chain in pre-BCR is an imperative for further B cell development [15].
BCR signaling is involved in the prevention of naïve B cell premature activation and expansion of autoreactive clones. At the same time, low-level “tonic” BCR signaling is essential for naïve B cell survival [16]. BCR antigen binding promotes mature B cell activation and further differentiation of naïve B cells via activation of PLC-γ2, PI3K/AKT and MAPK signaling pathways [17]. To ensure proper functionality of activated B cells, BCR signaling sustains survival, stimulates cell growth, and supports other related cellular adaptations via an array of signaling cascades including Ca2+ signaling, NF-κB activation, PI3K/AKT/mTOR, NFAT, ERK, and MAPK signaling [18]. Furthermore, BCR is vital for antigen presentation and subsequent T cell response activation and for B cell differentiation into antibody-producing plasma cells [19]. In particular, BCR-mediated antigen internalization is followed by intracellular antigen processing and subsequent surface presentation to CD4+ and CD8+ T cells [20]. BCR signaling also critically regulates activation-induced cytidine deaminase (AID)-mediated immunoglobulin class switch recombination [21,22]. Combination of activated BCR signaling with either a T cell-dependent (follicular T helpers) or T cell-independent (lipopolysaccharides or glycolipids) signal is crucial for B cell differentiation into antibody-secreting plasma cells or memory B cells [23]. BCR signaling driven plasma cell differentiation requires transcription factor Ets1 downregulation via Lyn-, PI3K-, BTK-, IKK2- and JNK-dependent pathways [24].
Additionally, BCR signaling contributes to the regulation of multiple other cellular processes in normal B cells including metabolism. For instance, it induces PI3K/AKT-dependent activation of glycolysis, oxidative phosphorylation, and glucose uptake [25,26]. BCR signaling also activates c-Myc with resulting enhancement of glycolysis and mitochondrial biogenesis [27]. BCR-initiated Ca2+ mobilization regulates metabolic reprogramming of naïve B cells which is required for their growth and further differentiation [18]. Maintenance of the balance between cellular growth and catabolic and anabolic processes is critical for correct B cell functionality and is primarily sustained via c-Myc and mTORC1 activity (which are both adjusted through BCR signaling) [28]. Importantly, BCR signaling was also implicated in metabolic regulation via autophagy upregulation [29]. BCR-mediated autophagy has been reported to be required for B cell activation [28].
Besides survival, activation, and proliferation, BCR signaling may prime B cells to anergy and cellular death to ensure B cell tolerance [30]. For instance, BCR signaling has been suggested to serve as a B cell quality control. Only moderate-intensity BCR signaling promotes positive selection, while BCR ligation downregulates BCR expression, reduces pro-survival PI3K/AKT signaling, and provides negative selection [31]. Inappropriately activated BCR may lead to B cell apoptosis [32]. BCR-mediated pro-apoptotic signaling has been associated with Ca2+-dependent and mitochondrial pathways [30].
Therefore, not only the type of BCR signaling, but also its intensity varies during B cell development and can determine the cell fate of B cells and their involvement in the immune response.
3. B-Cell Derived Malignancies and BCR Signaling
According to the recently updated 5th classification of lymphoid neoplasms (the World Health Organization Classification of Haematolymphoid Tumours), B-cell malignancies include the following categories: tumor-like lesions with B-cell predominance, precursor B-cell neoplasms (B lymphoblastic leukemias), mature B-cell neoplasms, and plasma cell neoplasms and other diseases with paraproteins [33]. Mature B-cell neoplasms include, e.g., pre-neoplastic and neoplastic small lymphocytic proliferations (e.g., chronic lymphocytic leukemia, CLL), multiple types of non-Hodgkin lymphomas (NHLs), and Hodgkin lymphomas [33]. The most common subtypes of NHL are diffuse large B-cell lymphoma (DLBCL) and follicular lymphoma (FL), diagnosed in approximately 25–30% and 20% of NHL patients, respectively [34,35,36,37]. In most cases, B-cell-derived tumors retain surface expression of BCR which variably supports malignant cell growth and survival [38,39]. BCR signaling has been shown to drive the growth and evolution of B-cell acute lymphoblastic leukemia (B-ALL), chronic lymphocytic leukemia (CLL), and multiple types of NHLs [12,40,41,42]. Pathogenic BCR signaling has been extensively studied and clearly demonstrated for DLBCL. Additionally, there is evidence that BCR supports tumor cell growth and survival in mantle cell lymphoma (MCL), FL, Burkitt’s lymphoma, and marginal zone lymphoma [43,44,45,46].
BCR signaling supports tumor cell growth and survival via various mechanisms. The first described tumorigenic mode of BCR signaling was the so-called “chronic active” BCR signaling triggered by self-antigen binding. Chronic active BCR signaling supports the viability and growth of malignant B cells mainly through the NF-κB signaling pathway [47,48]. Recently, it was shown that frequent lymphoma-associated mutations of MYD88 (myeloid differentiation primary response 88) adaptor protein lead to its spontaneous association with Toll-like receptor 9 (TLR9) and BCR, forming a My-T-BCR complex capable to trigger NF-κB activation [49]. Moreover, it was shown that lymphoma growth is also supported by antigen-independent, constitutive, lower intensity “tonic” BCR signaling. Tonic BCR signaling supports the growth and survival of tumor cells mostly via the PI3K/AKT/FOXO1 signaling pathway [50,51]. Antigen-independent cell autonomous BCR signaling with features of antigen-triggered BCR signaling was identified in CLL [40]. Importantly, in DLBCL, the type of BCR signaling (antigen driven or similar vs. tonic) reflects gene expression profiling-based cell-of-origin classification into the activated B cell like (ABC) DLBCL subtype and germinal center B cell like (GCB) DLBCL subtype, respectively [52].
Given the importance of BCR signaling in B-cell derived malignancies, its inhibition is one of the novel therapeutic approaches. It is represented mainly by three BTK (Bruton’s tyrosine kinase) inhibitors (ibrutinib, acalabrutinib, and zanubrutinib) approved and frequently used in the treatment of certain B-cell derived neoplasms. BTK inhibitors are effective; however, their toxicity and common resistance development represent substantial challenges that motivate the search for additional BCR signaling targeted inhibitors [53].
Important considerations regarding types of BCR signaling come from genomic studies, as documented in DLBCL. Distinct patterns of BCR signaling are reflected in tumor mutational patterns, which further expand the above-mentioned cell-of-origin DLBCL classification. Based on the spectrum of somatic alterations, genomic studies identified five to seven distinct genetic DLBCL subtypes [54,55,56,57,58]. The MCD (combined MYD88L265P and CD79B mutations), N1 (mutated NOTCH1), and A53 (aneuploid and TP53 inactivation) subtypes are significantly overlapping with the ABC DLBCL subtype, whereas EZB (mutated EZH2 and translocated BCL2), ST2 (mutated SGK1 and TET2), and BN2 (translocated BCL6 and mutated NOTCH2) are overlapping with GCB DLBCL [56]. Alternative classifications were published by Chapuy et al., including clusters 1 to 5 (BN2-DLBCL, A53-DLBCL, EZB-DLBCL, ST2-DLBCL, and MCD-DLBCL, respectively); and by Lacy et al., including MYD88, BCL2, SOCS1/SGK1, TET2/SGK1, and NOTCH2 clusters [55,58]. Pedrosa et al. later attempted to unite and simplify the existing classifications through the assessment of the mutational status of only 26 genes and BCL2 and BCL6 translocation status to facilitate their clinical implementation (two-step genetic DLBCL classifier; 2-S). The suggested 2-S subtypes are N12-S, EZB2-S, MCD2-S, BN22-S, and ST22-S [54]. Importantly, none of the above-mentioned genetic studies were able to assign all cases, leaving a substantial proportion of tumors unclassified. On the other hand, genetic studies provided insights into the contribution of CD79a and CD79b (and their mutations) towards tumorigenesis and lymphoma development [54,55,56,57,58].
4. Extracellular, Transmembrane, and Intracellular Domains of CD79a/CD79b Are Functionally Distinct in BCR
BCR organization has been intensively studied in recent years. Therefore, information about the BCR structure provided novel insights into molecular mechanisms of BCR signal initiation and signal transduction. In 2022, two independent groups successfully identified the human BCR structure using cryo-electron microscopy [59,60]. Furthermore, the identification of the murine BCR structure supplemented the data on human BCR [61].
Each of the approximately 120,000 BCR receptor molecules present at the surface of a mature B cell contains a membrane-embedded Ig (class M, D, G, A, or E) composed of two light and two heavy chains linked with disulfide bonds [61,62]. There is a strong evidence that the immunoglobulin-CD79a/CD79b stoichiometric ratio is 1:1, supporting the attachment of CD79a/CD79b to the immunoglobulin (in a non-covalent manner) [59,60,61,63,64]. The membrane attachment of the immunoglobulin molecule is mediated via C-terminal regions of Ig heavy chains, specifically by their transmembrane domains [65]. Multiple studies indicated that the cytoplasmic tails of membrane-bound immunoglobulins mediate isotype-specific BCR signaling. The immunoglobulin tail tyrosine (ITT) signaling motif was described in membrane-bound IgG and seems to mediate the reactivation of IgG-switched memory B cells. Following antigen-induced SYK (spleen tyrosine kinase)-mediated phosphorylation, the ITT facilitates docking of adaptor protein Grb2 (growth factor receptor-bound protein 2). Grb2 in turn recruits BTK, which leads to the amplification of Ca2+ mobilization and resulting amplification of BCR signaling [66,67,68,69]. However, the CD79a/CD79b heterodimer is critically important for a general BCR signal transduction [70,71]. CD79a/CD79b heterodimeric unit contains three domains (extracellular, transmembrane, and intracellular) and is bound together with disulfide bonds formed between cysteine residues of its extracellular domains [72]. Interestingly, CD79a and CD79b extracellular domains are the least conserved regions of CD79a and CD79b molecules with significant interspecies variability. Experiments in mouse B cells showed that their replacement with the human counterpart does not affect BCR assembly and signaling [73].
Interactions between Ig and CD79a/CD79b complex result in the formation of four-helix transmembrane bundle whose conserved nature assures correct assembly of any BCR isotype [60,74]. These tight interactions within the cell membrane are crucial for the assembly, stability and functionality of BCR, suggesting co-folding of BCR components during BCR complex formation [63]. Moreover, recent study demonstrated that the assembly and signaling features of BCR additionally depend on protein–protein interactions between transmembrane domains of CD79a and CD79b [72].
Specific interactions between the membrane-bound immunoglobulin and CD79a/CD79b are dependent on the BCR isotype. The IgM immunoglobulin binds to the CD79a/CD79b heterodimer extracellular domain from the side, whereas the IgG immunoglobulin interacts with the top of the CD79a/CD79b complex. It further suggests isotype specific differences in BCR signaling [74,75].
Intracellular domains of CD79a and CD79b have an immunoreceptor tyrosine-based activation motif (ITAM) made up of 26 amino acid residues and containing two critical tyrosine residues. ITAM tyrosines can undergo phosphorylation by the Src-family of kinases and are directly involved in BCR signal mediation [17,76,77]. Furthermore, it seems that ITAM phosphorylation also contributes to the regulation of BCR internalization [78].
The in-depth analysis of the cryo-electron microscopy BCR structure and additional studies of BCR functionality revealed that individual CD79a and CD79b domains mediate distinct processes to coordinate BCR signaling (Figure 1).
5. The CD79a/CD79b Heterodimer Is Critically Important for BCR Functionality
BCR plays a pivotal role in B cell biology. It regulates antigen uptake, antigen presentation, cell survival, proliferation, activation, differentiation, metabolism, B cell negative selection, cellular death, and B cell anergy [1,11,17,19,28,79,80,81,82,83]. Results of multiple studies and our current understanding of BCR signaling suggest that the CD79a/CD79b heterodimer is among key decisive regulators of BCR signaling and B cell fate.
CD79a and CD79b were implicated in the regulation of membrane-bound IgM expression and BCR complex formation and stability. The functional CD79a/CD79b complex promotes IgM transport and increases its surface levels via adjusting its glycosylation [2]. Furthermore, CD79a and CD79b facilitate assembly and steadiness of BCR by stabilizing each other to form the CD79a/CD79b dimer, mediate glycosylation of each other and IgM, as well as deliver the assembled receptor to the Golgi apparatus [2,84]. CD79a- and CD79b-mediated intracellular trafficking has been suggested to depend on their transmembrane domains’ interactions and ubiquitination patterns [2,78,85].
Two of the main pathways mediating survival signals downstream of BCR are NF-κB signaling (e.g., anti-apoptotic genes upregulation) and PI3K/AKT signaling (e.g., via its downstream effector transcriptional factor FOXO1) [86,87,88,89]. In naïve B cells, both CD79a and CD79b are required to physically interact with B-cell activating factor receptor (BAFFR), whose ligand, the B-cell activating factor (BAFF), initiates pro-survival PI3K/AKT signaling upon binding to BAFFR [90]. BCR signaling, combined with co-stimulation from CD40, BAFFR, and TLRs, regulates antibody-mediated immune responses requiring class switch recombination [21]. Interestingly, He et al. reported that association of CD19 with CD79b alone could trigger an alternative pathway to promote survival, fitness, and growth of B cells in a PI3K-dependent fashion [76]. Antigenic BCR stimulation and consequent signaling events also induce association of CD79a/CD79b heterodimers with MHC class II molecules. CD79a/CR79b heterodimers thus contribute to signal transduction following MHC class II activation (engaging Src-family tyrosine kinases and Ca2+ signal) [91,92]. The indirect mechanism by which the CD79a/CD79b heterodimer promotes cell survival is the already mentioned enhancement of IgM surface expression [2]. BCR signaling could also induce autophagy, which is involved in the regulation of cell survival and metabolic homeostasis of B cells [28,93]. Furthermore, B cell polarization and BCR intracellular trafficking are regulated by autophagy-related proteins mobilized upon BCR signaling activation [94]. However, the exact role of CD79a and CD79b in these signaling events, as well as details about their interaction with other BCR co-receptors, are not yet fully known and may not be limited to a simple recruitment of BCR signaling mediators. Importantly, autophagy-related regulation of BCR trafficking may complement the already reported effects of CD79a and/or CD79b phosphorylation and ubiquitination on antigen processing and presentation [78,85]. As mentioned above, proliferation and activation of B cells is supported by BCR-mediated PI3K signaling activation and consequent stimulation of mitochondrial biogenesis and elevated glucose uptake [26,28,95]. Nevertheless, it is important to emphasize that while BCR signaling is important for metabolic reprogramming of B cells, the ITAM-containing CD79a and CD79b BCR components are not indispensable for it [96].
The critical importance of CD79a and CD79b BCR subunits for BCR functionality, signal transmission, and signal regulation was largely established by studies of CD79a and CD79b posttranslational modifications.
6. BCR Signaling Is Regulated by Phosphorylation, Ubiquitination, and Glycosylation of CD79a and CD79b
ITAM phosphorylation is critical for signal transduction in ITAM-containing receptors. ITAM motifs are relatively common across various receptors in different immune cells including, e.g., T cells, NK cells, macrophages, or dendritic cells [97]. It is generally accepted that phosphorylation of both ITAM tyrosine residues by Src-family tyrosine kinases such as Lyn, Fyn, or Blk recruits SYK and the resulting complex activates downstream signaling pathways (e.g., PI3K/AKT, NF-κB, and MAPK) [17,38,98,99]. Furthermore, recent studies suggested that phosphorylation patterns of CD79a and CD79b ITAMs might affect BCR cellular localization and determine which downstream kinase is recruited to modulate BCR signaling (Figure 2).
Upon antigen binding, most cell surface BCR molecules are internalized for antigen processing and further presentation [5,100]. This process can occur even when ITAM tyrosine residues remain non-phosphorylated [78,101]. Importantly, non-phosphorylated ITAM-mediated BCR endocytosis is critically dependent on mCD79b Y195, but not mCD79a Y182 (murine CD79a, these tyrosine residues correspond to CD79b Y196 and CD79a Y188 in humans, respectively) and relies on adaptor protein 2 (AP2, an important mediator molecule involved in clathrin-mediated endocytosis). Phosphorylation of ITAM tyrosine residues prevents AP2 binding to the BCR YxxØ endocytosis motif and hence BCR internalization [101].
Results of multiple studies indicate that mono- or bis-phosphorylation of ITAM tyrosine residues determines whether Lyn or SYK is recruited. Lyn is recruited upon ITAM mono-phosphorylation, while SYK docking occurs in response to ITAM bis-phosphorylation (observed only in approximately 20% of activated BCR molecules) [102,103,104]. It has been also reported that Src-family kinases are preferentially recognized by CD79a in contrast to CD79b [105]. Comparing the two CD79a ITAM phosphorylation sites, mCD79a Y182 mediates Lyn recruitment in comparison to mCD79a Y193 (murine CD79a, these tyrosine residues correspond to Y188 and Y199 in human CD79a, respectively) [104]. Lyn acts as a molecular switch in BCR signaling, working in a bidirectional manner. Like other Src-family kinases, it phosphorylates CD79a/CD79b ITAMs to provide SH2 domain-associated SYK docking. On the other hand, Lyn phosphorylates ITIMs (immunoreceptor tyrosine-based inhibitory motifs) in ITIM containing receptors with consequent recruitment of phosphatases such as SHIP-1 (Src homology region 2 domain-containing inositol 5′ phosphatase 1), SHP-1 (Src homology region 2 domain-containing tyrosine phosphatase), and PTEN (phosphatase and tensin homolog) to downregulate BCR signaling [102,106,107]. Lyn-mediated SYK recruitment activates the B-cell linker (BLNK) protein, which promotes assembly of phosphoinositide phospholipase C-γ-2 (PLC-γ2), BTK, and adaptor protein Grb2 into a multimolecular regulatory complex leading to downstream NF-κB and PI3K/AKT signaling activation [17]. However, within components of this classical BCR signalosome, Lyn is not indispensable. Lyn deficiency in Lyn knock out (KO) models can be compensated by other members of Src-family kinases [108,109]. At the same time, Lyn is critical for signalosome-independent IL-4-mediated alternative pathway of BCR signaling activation [108].
The above-mentioned Lyn-dependent negative regulation of BCR signaling relies on SHIP-1- and PTEN-catalyzed dephosphorylation of phosphatidylinositol (3,4,5)-trisphosphate (PtdIns(3,4,5)P3), blocking the PI3K/AKT pathway [103]. Moreover, Lyn-mediated SHP-1 activation leads to CD79a/CD79b ITAMs and SYK dephosphorylation, forming a negative feedback loop [110,111]. Therefore, Lyn determines the signal transduction strength, mediating the balance between phosphorylation/dephosphorylation of key regulatory intracellular BCR signaling components. Therefore, it affects B cell fate decisions. Lyn recruitment, which depends on phosphorylation patterns of CD79a/CD79b, upregulates intracellular Ca2+ signaling through SYK-mediated PLC-γ2 activation and downregulates it via PtdIns(3,4,5)P3 degradation (mediated by SHIP-1 and PTEN). It was reported that quantitatively distinct Ca2+ signaling patterns downstream of BCR activation regulate B cell survival by NF-κB engagement and B cell proliferation via NFAT, mTORC1, and c-Myc activation [18].
Experimental evidence indicates that other than classical ITAM tyrosines of CD79a (amino acid positions 188 and 199) are also substrates of phosphorylation and are involved in regulation of BCR signaling. mCD79a Y176 and Y204 are required for BLNK recruitment (a key adaptor phosphorylated by SYK). mCD79a Y204 phosphorylation is of particular importance for BLNK recruitment. It places SYK and BLNK in a close proximity to allow further BLNK activation. It also links SYK with downstream BCR signaling events including Ca2+ mobilization, NF-κB, ERK, and JNK activation [112,113,114]. The murine mCD79a Y176 corresponds to human CD79a Y182 (a third tyrosine within the ITAM domain) and murine mCD79a Y204 corresponds to human CD79a Y210 (a non-ITAM downstream tyrosine). Therefore, both ITAM and non-ITAM tyrosine residues are subject to phosphorylation and are all crucial for BCR signal transduction and regulation of its strength. The strength of BCR signaling also depends on intricate equilibrium between kinases and phosphatases.
Phosphorylation is certainly a key covalent post-translational modification of CD79a/CD79b which regulates BCR signaling. However, several studies suggested that both CD79a and CD79b lysine sites are utilized for ubiquitin attachment and are therefore prone to ubiquitination [78,85,115,116,117,118]. As mentioned previously (Figure 1), ubiquitination sites are located in the intracellular domain of CD79a and CD79b [115]. Upon BCR-antigen binding, CD79a and CD79b are subject to rapid ubiquitination (within 5 min) [117]. Time-dependent kinetics of CD79a and CD79b ubiquitination and its role in BCR receptor function still needs to be elucidated, but it has been demonstrated that CD79b is ubiquitinated first, followed by a CD79a ubiquitination [85].
Ubiquitination might have a much wider significance in proximal BCR signaling regulation in general since CD79a and CD79b are ubiquitinated concurrently with Lyn and SYK. Importantly, CD79a phosphorylation (but not CD79b phosphorylation) seems to promote CD79a ubiquitination. Moreover, concurrent activation of SYK employs ubiquitin ligases such as c-Cbl and Cbl-b, which may further recruit Itch (another ligase involved in ubiquitination-dependent BCR signal regulation) [117,119]. Involvement of ubiquitination at early stages of BCR signaling suggests that it may be a versatile signaling mark. Several studies showed that CD79a and/or CD79b ubiquitination specifically targets BCR for endocytosis even if ITAM tyrosines are phosphorylated and that it affects intracellular trafficking, endosome signaling, and antigen processing and presentation [78,85,116,118]. Veselits et al. reported that CD79b ubiquitination is necessary for PI3K activation and PIP3 accumulation in BCR-containing endosomes, affecting consequent sorting into the major histocompatibility complex (MHC) class II antigen-presenting compartment and endocytic trafficking [116]. Moreover, KLHL14 (Kelch-Like Family Member 14, a tumor-suppressing chaperone regulating protein folding, frequently mutated in lymphomas) promotes CD79a and CD79b ubiquitination and their consequent downregulation [115]. Since CD79a/CD79b ubiquitination is involved in many aspects of BCR signaling, it could be potentially targetable; however, the exact mechanisms of CD79a/CD79b ubiquitination and its regulation are not fully clarified.
Recent studies also showed that CD79a/CD79b could be N-glycosylated and have linked this modification to the regulation of BCR surface expression. Abnormal glycosylation of CD79a results in retention of BCR complex in the endoplasmic reticulum for further proper folding and assembly, leading to a reduction in its translocation to the cell membrane [84]. Apart from the above-mentioned CD79a/CD79b ubiquitination, KLHL14 protein affects also CD79a/CD79b glycosylation. It regulates immature glycosylated BCR stability and turnover in the endoplasmic reticulum [115].
In summary, post-translational modifications of CD79a/CD79b heterodimer define its own assembly, BCR expression, BCR endocytosis, strength of BCR signaling, intracellular trafficking, and antigen processing and presentation.
7. BCR Signaling Is Regulated at the Level of CD79a/CD79b Heterodimer by Physical Interactions with Regulatory Molecules
As already mentioned above, CD79a/CD79b heterodimer or its components can physically interact with a wide range of regulators to modulate BCR signaling (summarized in Figure 3). Classical BCR signaling requires direct Lyn recruitment or SYK docking to promote further activation of downstream kinases [102,103,104]. On the other hand, multiple studies have recently demonstrated that CD79a and CD79b might have substantially larger interactomes.
As mentioned above, CD79a/CD79b heterodimeric unit determines BCR internalization via its ubiquitination and ITAM phosphorylation. Mono-phosphorylation of CD79 ITAMs recruits Lyn, while bis-phosphorylation docks SYK with further activation of BTK and PLCγ2 and resulting downstream activation of NF-κB and PI3K/AKT signaling. CD79a/CD79b unit mediates also co-signaling from CD19, BAFFR, MHC II, and TLR9 and is regulated by KLHL14 and TRAF3 (tumor necrosis factor receptor (TNFR)-associated factor 3). CD79b seems to localize in a close proximity to CD19 even when it is not associated with other components of BCR (i.e., Ig and CD79a). This interaction helps to maintain pro-survival signal in B cells in an ITAM/PI3K-dependent manner [76]. PI3K recruitment upon stimulation of CD19 co-receptor requires its Lyn-dependent phosphorylation [120]. Direct interactions of CD79a and CD79b with TLR9 lead to MYD88 recruitment and hence NF-κB signaling upregulation via the MYD88-TLR9-BCR supercomplex [49]. In addition to NF-κB signaling, ligand binding to BAFFR (which could be either attached or located closely to CD79a and CD79b) upregulates also PI3K/AKT as another critical BCR downstream pathway [90]. Furthermore, CD79b, along with other proteins involved in proximal BCR signaling (SYK and BTK), interacts with TRAF3 (a negative regulator of BCR signaling). Therefore, this interaction results in reduction of non-canonical NF-κB and MAPK/ERK pathway activation without affecting canonical NF-κB and PI3K/AKT signal [121]. Notably, TRAF3 has been well established as a regulator of CD40 and BAFFR signaling in B cells [122,123]. The above-mentioned KLHL14 is another negative regulator of BCR signaling which physically interacts with CD79a and CD79b [115]. As mentioned above, CD79a/CR79b heterodimers could also associate with MHC class II and contribute to its downstream signal transduction [91,92].
Expansion of CD79a and CD79b molecular interaction information further supports the critical role of these BCR components in a complex regulation of downstream BCR signaling. It can be assumed that CD79a/CD79b unit, especially its CD79b component, may represent a signaling hub mediating crosstalk between BCR and its co-receptors.
8. CD79a and CD79b Are Important Regulators of Proximal and Distal BCR Signaling in Malignant B Cells
Availability of next-generation sequencing has significantly expanded our understanding of mutational profiles in B-cell derived malignancies, increased the number of known tumor driver genes, and provided information about additional mechanisms of tumorigenesis. Tumorigenic contribution of CD79A and CD79B mutations is well studied and could be demonstrated in NHL as an example. Mutation frequency of CD79B varies between 4 and 23%, largely depending on NHL type [54,57,124,125]. Mutations of CD79A are present in approximately 3–4% of NHL [57,58]. CD79A and CD79B mutations most frequently affect ITAM regions. In general, CD79A alterations primarily result in removal of the entire ITAM region, while CD79B mutations frequently affect the first tyrosine residue of ITAM, the Y196 [41,126,127]. The main example of uneven distribution of CD79A and CD79B mutations between lymphoma subtypes is DLBCL. CD79A and CD79B mutations are observed in up to 30% of ABC DLBCL cases and only in 3% of GCB DLBCL tumors [41,48,57,126,128,129]. None of the currently proposed genetic clustering algorithms uses CD79A mutations as a genetic feature of any DLBCL genetic subtype. However, frequency of CD79A alterations is higher in EZB cluster [54,55,56,57,58,130]. In contrast to CD79A, CD79B mutations are one of the most enriched genetic features of MCD cluster and are very frequent in BN2 and A53 clusters in the GenClass and LymphGen algorithm-based genetic classifications, respectively [56]. According to the alternative genetic DLBCL classifications, CD79B mutations are enriched in Cluster 5, MYD88, and MCD2-S genetic subtypes, which are all MCD cluster equivalents [54,55,58]. This genetic DLBCL subtype (with high frequency of CD79B mutations) is robust and omnipresent across all genomic classifications of DLBCL and is strongly associated with active BCR signaling. In lymphoma, CD79B mutations augment BCR surface levels via reduction of BCR endocytosis, preventing BCR binding to clathrin-coated pits [3,41,48,101,126,131]. Importantly, BCR signaling strength seems to be proportional to CD79b expression levels in NHL [132]. Another widely recognized mechanism through which mutated CD79b increases BCR signaling is its inability to properly activate Lyn, a Src-family tyrosine kinase triggering negative feedback loop-based inhibition of BCR signaling [3,41,126,128,131]. Reduced Lyn activity also contributes to overcoming B cell anergy [41]. Importantly, CD79b with mutated ITAMs promotes BCR clustering in ABC DLBCL, which is a characteristic consequence of BCR-antigen binding [3,38,50,133]. It indirectly supports the important role of CD79b protein alterations in chronic active BCR signaling augmentation. Therefore, CD79b mutations might promote pro-survival active BCR signaling via BCR surface levels upregulation, enhancement of BCR clustering, inhibition of BCR internalization, and reduced BCR signaling negative regulation. On top of ITAM tyrosines mutations, Andrades et al. reported that CD79B gene is frequently affected by recurrent splice site mutations in DLBCL. These mutations frequently lead to intron 4 retention with premature termination of CD79b translation. It also results in BCR overexpression and enhancement of NF-κB and AKT signaling [134]. CD79a and CD79b molecules (and possibly their mutations) further affect the CD79b-CD19 and BCR-BAFFR mediated PI3K/AKT activation and BCR/TLR9/MYD88 and CARD11/BCL10/MALT1 complexes formation with consequent NF-κB signaling activation.
CD79A mutations are much less frequent and were not identified within any genetic DLBCL cluster. On the other hand, it was reported that CD79A mutations can contribute to activation and enhancement of chronic active BCR signaling in ABC DLBCL as well [135]. Importantly, wt CD79a, specifically its ITAM Y188 and its phosphorylation, are critical for tonic BCR signal mediation in GCB DLBCL [50].
Frequencies and signaling consequences of CD79A and CD79B mutations are overviewed in Table 1 and Table 2, respectively. Most genetic studies focused only on ITAM regions, therefore, data on frequencies of non-ITAM region mutations are limited. However, Lohr et al. reported that in DLBLC, 12.5% of CD79B mutations affect non-ITAM regions [136]. High rate of mutations in extracellular and transmembrane domains of CD79b (over 30% of patients) was detected also in CLL [137]. Therefore, it would be of great interest to analyze specifically frequency and eventual signaling consequences of mutations affecting non-ITAM regions across various B-cell derived malignancies.
Taken together, it could be hypothesized that chronic active BCR signaling in ABC DLBCL relies primarily on CD79b, while CD79a contributes more to tonic BCR signaling in GCB DLBCL. However, more studies are necessary to elucidate the exact contribution of either CD79a or CD79b (and their mutations) to various types and states of BCR signaling in lymphoma. An overview of CD79a/CD79b involvement in lymphoma associated BCR signaling is provided in Figure 4.
Importantly, ABC DLBCL tumor cell survival is critically dependent on BCR triggered NF-κB activation [50,154,155,156]. BCR downstream NF-κB activation is mediated by CD79a and CD79b and is enhanced by their mutations. However, within the pathologically active BCR signaling in lymphoma, CD79A or CD79B mutations cannot be considered separately from other mutations. It is only one molecular event within a quite complex BCR signaling and NF-κB activation process. As an example of this complexity, CD79A and CD79B mutations were included in a large network of 153 lymphoma-altered genes related to NF-κB signaling [124]. Other studies also reported that CD79A and CD79B mutations modify distal BCR signaling in collaboration with other mutations of the BCR signaling network via modulation of NF-κB signaling (with an effect on survival, apoptosis, and proliferation of tumor cells). Mutated CD79a or CD79b proteins cannot trigger BCR signaling by itself; they “only” amplify the signal and modulates its strength and direction [135]. This relationship was described specifically for CARD11 (caspase recruitment domain family member 11) mutations in ABC DLBCL. CARD11 acts as a component of the CARD11-BCL10-MALT1 signalosome complex involved in bridging proximal and distal BCR signaling towards NF-κB activation [157,158]. CARD11 mutations seem to be necessary for mutated CD79a/CD79b-mediated NF-κB activation [48]. In addition, viability of mutated CD79B ABC DLBCL cells critically relies on BTK that fuels canonical NF-κB signaling via regulation of downstream mucosa-associated lymphoid tissue lymphoma translocation protein 1 (MALT1) [129]. CD79B mutations not only sustain anti-apoptotic and proliferation-inducing NF-κB signaling directly but also support the interplay between other BCR-associated signaling pathways to ensure the survival of ABC DLBCL cells. It was reported that in two model ABC DLBCL cell lines with mutated CD79B, PI3K signaling and downstream PDK1 (putative 3-phosphoinositide-dependent kinase 1) activation are essential events for CARD11-BCL10-MALT1 signalosome complex-mediated NF-κB activation [159]. This suggests a substantial interaction between PI3K signaling and the NF-κB pathway in CD79B-mutated ABC DLBCL. In MCL, pro-survival PI3K/AKT/mTOR signaling is sustained by wt CD79a and requires activity of Lyn [160]. This is consistent with the role of CD79a in promoting pro-survival PI3K/AKT/mTOR signaling in GCB DLBCL [50]. In contrast to ABC DLBCL, GCB DLBCL cells rely on the PI3K/AKT/mTOR to support their survival [161,162]. Therefore, GCB DLBCL is characterized by tonic BCR signaling where CD79a phosphorylation results in activation of downstream PI3K/AKT pathway [50].
In lymphomas, CD79B mutations also commonly co-occur with MYD88 mutations, especially MYD88L265P. MYD88 mutations have higher prevalence in cases with extranodal localization and mediate active BCR signaling phenotype within the MCD genotype of DLBCL [3,56,58,126,127,130,163,164,165]. Multiple studies reported a combined biological effect of CD79B and MYD88 mutations. Importantly, none of these two mutated genes can induce malignant transformation alone as they need to act in collaboration with other driver mutations [127,166]. It has been shown that only the combination of CD79B and MYD88 mutations prevented anergy of autoantigen-stimulated B cells and allowed their plasmablastic differentiation [152]. The most common MYD88L265P mutation results in overactivation of MYD88 protein, a molecular adaptor that normally promotes NF-κB signaling via IRAK (IL-1 receptor-associated kinase) recruitment and formation of the so-called myddosome complex [167]. Myddosomes are large oligomeric signaling complexes that are assembled within TLRs and IL-1 signaling. Myddosomes are primarily formed by MYD88 and IRAKs and act as a scaffold for further downstream signal transduction [168,169,170,171]. Phelan et al. demonstrated that CD79B-MYD88 mutations simultaneously facilitate intermolecular interactions to form a three-component MYD88-TLR9-BCR (My-T-BCR) supercomplex, which drives NF-κB and mTOR signaling activation [49]. Furthermore, lymphoma sequencing studies revealed that the network of concurrently mutated genes in MCD DLBCL cluster includes inactivating mutations of KLHL14, which promotes My-T-BCR-dependent NF-κB signaling via reduction of CD79a and CD79b ubiquitination [115]. It is worth mentioning that enhancement of CD79b-mediated non-canonical NF-κB signaling has been associated with TRAF3 insufficiency in lymphoma [121].
In addition to the My-T-BCR supercomplex, CD79a/CD79b maintains pro-survival PI3K/AKT signaling via interaction with BCR co-receptors BAFFR and CD19 in B-cell NHL [76,90]. The pro-survival signal comes from BAFF-dependent Lyn-mediated activation of PI3K/AKT [90]. A similar PI3K-dependent pro-survival signal is sustained by CD79b through CD19 activation. It is ITAM-dependent, can occur even in the absence of other BCR components (IgM and CD79a), and is considered an alternative lymphoma survival supporting mechanism [76].
Taken together, CD79a and CD79b BCR components could be considered critical regulators of altered signal flow in the complex network of deregulated BCR signaling in NHL. This suggests that CD79a/CD79b complex might represent a suitable and universal therapeutical target for B-cell malignancies and possibly other B-cell-related disorders. On the other hand, BCR signaling deregulation occurs in response to multiple mutations in several interacting and BCR regulatory elements adjusting pro-survival BCR signaling in a relatively diverse manner.
9. Open Questions and Future Directions
Next-generation sequencing-based subtyping of DLBCL has improved the accuracy of disease outcome prediction. Particularly, concomitant CD79B and MYD88 mutations (characteristic for the MCD class of DLBCL) are associated with worse prognosis [3,55,57,126,172,173]. Importantly, a recent DLBCL meta-analysis showed that CD79B mutations had higher predictive value for disease progression and treatment outcomes than MYD88 mutations [164]. Inferior survival of patients with CD79B mutations might be related to a specific disease biology and associated with an increased risk of chemo-refractory disease and relapse [54]. Indeed, transcriptomic, and proteomic studies showed downregulation of CD79b in refractory DLBCL. This suggests that CD79b targeted therapy, such as polatuzumab vedotin (anti-CD79b monomethyl auristatin E antibody-drug conjugate), might not be best suited in this clinical situation [174]. On the other hand, overexpression of CD79b in ABC DLBCL contributes to ibrutinib (a potent BTK inhibitor) resistance. Similarly, MCL tumors with higher baseline levels of CD79b expression required higher concentrations of ibrutinib for efficient BCR signal suppression [132,175,176]. None of these correlations were observed for CD79a upregulation. On the other hand, it was reported that BTK inhibition increases CD79a phosphorylation as a part of compensation feedback loop. It leads to activation of multiple key BCR signal mediators with consequent proximal BCR signal rewiring [177]. Increased CD79a/CD79b activity might be one of general resistance mechanisms. It was reported that CD79a-mediated Lyn-dependent activation of PI3K/AKT/mTOR signaling also occurs in bortezomib (an FDA-approved 26S proteasome inhibitor) resistant MCL [160]. Future studies regarding CD79b activation and interactions in malignant B cells are necessary to fully understand its contribution to drug resistance and to confirm its eventual diagnostic significance as a reliable resistance biomarker. Pre-clinical in vitro-based studies could provide general information how CD79a and CD79b altered expression, mutations, and/or phosphorylation patterns affect sensitivity to various inhibitors. The variability of CD79b expression levels between different types of NHLs and individual tumors (which likely predicts BCR activity) indicates that CD79b expression could be evaluated as a possible biomarker for effective BCR inhibition. The evaluation of CD79a and CD79b pre-treatment expression levels (as well as its treatment resistance-related changes) should be implemented in clinical trials with BCR signaling inhibitors. Moreover, CD79b assessment could be considered in studies of other targeted therapies where BCR signaling activation might contribute to treatment resistance. Explanation of CD79a/CD79b signaling details, as outlined below, might help to identify additional therapeutic options for personalized B-cell derived malignancies treatment.
BCR signaling, as well as the functional involvement of CD79a and CD79b molecules, has already been extensively studied; however, many important issues remain to be addressed. What are the underlying transcription factors, pathways, or other mechanisms regulating CD79a and CD79b expression? Implementation of genome-wide KO screen approaches in model lymphoma cell lines might help to identify these factors. It could also explain differences in CD79a and CD79b expression levels (in normal as well as malignant B cells) and lead to the identification of novel approaches for BCR signaling inhibition.
Multiple questions also remain to be answered in relation to CD79a/CD79b ITAM phosphorylation. How is the ITAM phosphorylation regulated? Do CD79a/CD79b conformational changes mediate CD79a/CD79b ITAM phosphorylation? What are the exact and detailed characteristics of CD79a/CD79b interactions with Src-family kinases? Src-family tyrosine kinases generally have a BCR signaling activation role; however, Lyn forms a negative feedback loop highlighting its unique and important role within BCR signal regulation. The complexity, interaction, activation/inhibition balance, and co-regulation between CD79a/CD79b and Src-family kinases are not fully clarified. Moreover, are there any other kinases and phosphatases involved in ITAM phosphorylation and dephosphorylation? Basic molecular biology studies using targeted KO, protein–protein interaction assessment, or structural biology methods might bring such information.
Further studies should also focus on mechanisms of CD79a and CD79b ubiquitination and glycosylation as another layer of BCR signal regulation. Similarly, it is important to identify how is BCR signaling affected by tumor-related disruption of redox homeostasis and reactive oxygen species (ROS) production [178,179]. Novel proteomics-based studies of the CD79a/CD79b interactome might provide additional information on these unresolved issues.
Given the heterogenous nature of B-cell derived malignancies, it is similarly important to identify mechanisms by which CD79A and CD79B mutations modulate BCR signaling balance towards a particular effector pathway or pathways in a complex landscape of multiple co-occurring driver mutations. The direct involvement of CD79a/CD79b in CD19, BAFFR and TLR9 signaling raises a question of their possible critical role in the formation of a mutual BCR-centered signaling network [49,76,90,180]. Is the CD79a/CD79b heterodimer a signaling hub coordinating signals from various BCR co-receptors? Interpretation of genomic data and experimental studies also suggest that CD79a and CD79b signal mediation might be substantially different between normal and malignant B cells.
10. Conclusions
CD79a and CD79b molecules are at the center of a very complex BCR signaling network with critical functional implications for normal as well as malignant B cells. Moreover, CD79a/CD79b unit mediates signaling crosstalk between BCR and its co-receptors, which supports its role as an important B cell signaling hub. Solving above outlined open issues could further expand our understanding of BCR signal initiation and propagation and allow us to further specify the complexity of CD79a/CD79b signaling and possibly identify novel therapeutic targets for B-cell related disorders.
Author Contributions
Conceptualization, O.H.; writing—original draft preparation, A.T.; writing—review and editing, K.K. and O.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Czech Health Research Council (AZV NV18-03-00117), Charles University, Prague (PRIMUS/17/MED/9, UNCE/MED/016, Cooperatio), project National Institute for Cancer Research (reg. No. LX22NPO5102)—funded by the European Union—Next Generation EU, Programme EXCELES, and by the EHA Ukraine Bridge Funding awarded by the European Hematology Association.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tanaka, S.; Baba, Y. B Cell Receptor Signaling. Adv. Exp. Med. Biol. 2020, 1254, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Huse, K.; Bai, B.; Hilden, V.I.; Bollum, L.K.; Våtsveen, T.K.; Munthe, L.A.; Smeland, E.B.; Irish, J.M.; Wälchli, S.; Myklebust, J.H. Mechanism of CD79A and CD79B Support for IgM+ B Cell Fitness through B Cell Receptor Surface Expression. J. Immunol. 2022, 209, 2042–2053. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhou, D.; Wang, L.; Zhu, L.; Ye, X. MYD88(L265P) and CD79B double mutations type (MCD type) of diffuse large B-cell lymphoma: Mechanism, clinical characteristics, and targeted therapy. Ther. Adv. Hematol. 2022, 13, 20406207211072839. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, Y.; Zhang, S.; Haneef, K.; Liu, W. Structural and immunogenomic insights into B-cell receptor activation. J. Genet. Genom. 2020, 47, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Avalos, A.M.; Ploegh, H.L. Early BCR Events and Antigen Capture, Processing, and Loading on MHC Class II on B Cells. Front. Immunol. 2014, 5, 92. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, L.; Sasaki, Y.; Calado, D.P.; Zhang, B.; Paik, J.H.; DePinho, R.A.; Kutok, J.L.; Kearney, J.F.; Otipoby, K.L.; Rajewsky, K. PI3 kinase signals BCR-dependent mature B cell survival. Cell 2009, 139, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.A.; Wiestner, A. Targeting B cell receptor signalling in cancer: Preclinical and clinical advances. Nat. Rev. Cancer 2018, 18, 148–167. [Google Scholar] [CrossRef]
- Young, R.M.; Staudt, L.M. Targeting pathological B cell receptor signalling in lymphoid malignancies. Nat. Rev. Drug Discov. 2013, 12, 229–243. [Google Scholar] [CrossRef]
- Puri, K.D.; Di Paolo, J.A.; Gold, M.R. B-cell receptor signaling inhibitors for treatment of autoimmune inflammatory diseases and B-cell malignancies. Int. Rev. Immunol. 2013, 32, 397–427. [Google Scholar] [CrossRef]
- Rawlings, D.J.; Metzler, G.; Wray-Dutra, M.; Jackson, S.W. Altered B cell signalling in autoimmunity. Nat. Rev. Immunol. 2017, 17, 421–436. [Google Scholar] [CrossRef]
- Liu, W.; Tolar, P.; Song, W.; Kim, T.J. Editorial: BCR Signaling and B Cell Activation. Front. Immunol. 2020, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Köhrer, S.; Havranek, O.; Seyfried, F.; Hurtz, C.; Coffey, G.P.; Kim, E.; Ten Hacken, E.; Jäger, U.; Vanura, K.; O’Brien, S.; et al. Pre-BCR signaling in precursor B-cell acute lymphoblastic leukemia regulates PI3K/AKT, FOXO1 and MYC, and can be targeted by SYK inhibition. Leukemia 2016, 30, 1246–1254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Srivastava, G.; Lu, L. The pre-B cell receptor and its function during B cell development. Cell Mol. Immunol. 2004, 1, 89–94. [Google Scholar] [PubMed]
- Winkler, T.H.; Mårtensson, I.-L. The Role of the Pre-B Cell Receptor in B Cell Development, Repertoire Selection, and Tolerance. Front. Immunol. 2018, 9, 2423. [Google Scholar] [CrossRef] [PubMed]
- Keren, Z.; Melamed, D. Antigen receptor signaling competence and the determination of B cell fate in B-lymphopoiesis. Histol. Histopathol. 2005, 20, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Neys, S.F.H.; Heukels, P.; van Hulst, J.A.C.; Rip, J.; Wijsenbeek, M.S.; Hendriks, R.W.; Corneth, O.B.J. Aberrant B Cell Receptor Signaling in Naïve B Cells from Patients with Idiopathic Pulmonary Fibrosis. Cells 2021, 10, 1321. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Jing, Y.; Yang, L.; Kang, D.; Jiang, P.; Li, N.; Cheng, J.; Li, J.; Li, X.; Peng, Z.; et al. The regulators of BCR signaling during B cell activation. Blood Sci. 2019, 1, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Berry, C.T.; Liu, X.; Myles, A.; Nandi, S.; Chen, Y.H.; Hershberg, U.; Brodsky, I.E.; Cancro, M.P.; Lengner, C.J.; May, M.J.; et al. BCR-Induced Ca2+ Signals Dynamically Tune Survival, Metabolic Reprogramming, and Proliferation of Naive B Cells. Cell Rep. 2020, 31, 107474. [Google Scholar] [CrossRef]
- McShane, A.N.; Malinova, D. The Ins and Outs of Antigen Uptake in B cells. Front. Immunol. 2022, 13, 892169. [Google Scholar] [CrossRef]
- Rastogi, I.; Jeon, D.; Moseman, J.E.; Muralidhar, A.; Potluri, H.K.; McNeel, D.G. Role of B cells as antigen presenting cells. Front. Immunol. 2022, 13, 954936. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, J.H. How the Signaling Crosstalk of B Cell Receptor (BCR) and Co-Receptors Regulates Antibody Class Switch Recombination: A New Perspective of Checkpoints of BCR Signaling. Front. Immunol. 2021, 12, 663443. [Google Scholar] [CrossRef] [PubMed]
- Vlachiotis, S.; Abolhassani, H. Transcriptional regulation of B cell class-switch recombination: The role in development of noninfectious complications. Expert Rev. Clin. Immunol. 2022, 18, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Tsai, D.-Y.; Hung, K.-H.; Chang, C.-W.; Lin, K.-I. Regulatory mechanisms of B cell responses and the implication in B cell-related diseases. J. Biomed. Sci. 2019, 26, 64. [Google Scholar] [CrossRef]
- Luo, W.; Mayeux, J.; Gutierrez, T.; Russell, L.; Getahun, A.; Müller, J.; Tedder, T.; Parnes, J.; Rickert, R.; Nitschke, L.; et al. A balance between B cell receptor and inhibitory receptor signaling controls plasma cell differentiation by maintaining optimal Ets1 levels. J. Immunol. 2014, 193, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Kluckova, K.; D’Avola, A.; Riches, J.C. Advances in Understanding of Metabolism of B-Cell Lymphoma: Implications for Therapy. Cancers 2022, 14, 5552. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.A.; Bleiman, B.F.; Wagner, D.J.; Dufort, F.J.; Mataraza, J.M.; Roberts, M.F.; Chiles, T.C. Antigen receptor-mediated changes in glucose metabolism in B lymphocytes: Role of phosphatidylinositol 3-kinase signaling in the glycolytic control of growth. Blood 2006, 107, 4458–4465. [Google Scholar] [CrossRef]
- Iperi, C.; Bordron, A.; Dueymes, M.; Pers, J.-O.; Jamin, C. Metabolic Program of Regulatory B Lymphocytes and Influence in the Control of Malignant and Autoimmune Situations. Front. Immunol. 2021, 12, 735463. [Google Scholar] [CrossRef]
- Raza, I.G.A.; Clarke, A.J. B Cell Metabolism and Autophagy in Autoimmunity. Front. Immunol. 2021, 12, 681105. [Google Scholar] [CrossRef]
- Watanabe, K.; Tsubata, T. Autophagy connects antigen receptor signaling to costimulatory signaling in B lymphocytes. Autophagy 2009, 5, 108–110. [Google Scholar] [CrossRef]
- Eeva, J.; Pelkonen, J. Mechanisms of B cell receptor induced apoptosis. Apoptosis 2004, 9, 525–531. [Google Scholar] [CrossRef]
- Nemazee, D. Mechanisms of central tolerance for B cells. Nat. Rev. Immunol. 2017, 17, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Yam-Puc, J.C.; Zhang, L.; Maqueda-Alfaro, R.A.; Garcia-Ibanez, L.; Zhang, Y.; Davies, J.; Senis, Y.A.; Snaith, M.; Toellner, K.-M. Enhanced BCR signaling inflicts early plasmablast and germinal center B cell death. iScience 2021, 24, 102038. [Google Scholar] [CrossRef] [PubMed]
- Alaggio, R.; Amador, C.; Anagnostopoulos, I.; Attygalle, A.D.; Araujo, I.B.O.; Berti, E.; Bhagat, G.; Borges, A.M.; Boyer, D.; Calaminici, M.; et al. The 5th edition of the World Health Organization Classification of Haematolymphoid Tumours: Lymphoid Neoplasms. Leukemia 2022, 36, 1720–1748. [Google Scholar] [CrossRef] [PubMed]
- Susanibar-Adaniya, S.; Barta, S.K. 2021 Update on Diffuse large B cell lymphoma: A review of current data and potential applications on risk stratification and management. Am. J. Hematol. 2021, 96, 617–629. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Barta, S.K. Diffuse large B-cell lymphoma: 2019 update on diagnosis, risk stratification, and treatment. Am. J. Hematol. 2019, 94, 604–616. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Garg, V.; Mallick, S.; Gogia, A. Current trends in diagnosis and management of follicular lymphoma. Am. J. Blood Res. 2022, 12, 105–124. [Google Scholar] [PubMed]
- Carbone, A.; Roulland, S.; Gloghini, A.; Younes, A.; von Keudell, G.; López-Guillermo, A.; Fitzgibbon, J. Follicular lymphoma. Nat. Rev. Dis. Primers 2019, 5, 83. [Google Scholar] [CrossRef]
- Profitós-Pelejà, N.; Santos, J.C.; Marín-Niebla, A.; Roué, G.; Ribeiro, M.L. Regulation of B-Cell Receptor Signaling and Its Therapeutic Relevance in Aggressive B-Cell Lymphomas. Cancers 2022, 14, 860. [Google Scholar] [CrossRef]
- Niemann, C.U.; Wiestner, A. B-cell receptor signaling as a driver of lymphoma development and evolution. Semin. Cancer Biol. 2013, 23, 410–421. [Google Scholar] [CrossRef]
- Dühren-von Minden, M.; Übelhart, R.; Schneider, D.; Wossning, T.; Bach, M.P.; Buchner, M.; Hofmann, D.; Surova, E.; Follo, M.; Köhler, F.; et al. Chronic lymphocytic leukaemia is driven by antigen-independent cell-autonomous signalling. Nature 2012, 489, 309–312. [Google Scholar] [CrossRef]
- Young, R.M.; Phelan, J.D.; Wilson, W.H.; Staudt, L.M. Pathogenic B-cell receptor signaling in lymphoid malignancies: New insights to improve treatment. Immunol. Rev. 2019, 291, 190–213. [Google Scholar] [CrossRef] [PubMed]
- Valla, K.; Flowers, C.R.; Koff, J.L. Targeting the B cell receptor pathway in non-Hodgkin lymphoma. Expert Opin. Investig. Drugs 2018, 27, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Fichtner, M.; Dreyling, M.; Binder, M.; Trepel, M. The role of B cell antigen receptors in mantle cell lymphoma. J. Hematol. Oncol. 2017, 10, 164. [Google Scholar] [CrossRef] [PubMed]
- Sachen, K.L.; Strohman, M.J.; Singletary, J.; Alizadeh, A.A.; Kattah, N.H.; Lossos, C.; Mellins, E.D.; Levy, S.; Levy, R. Self-antigen recognition by follicular lymphoma B-cell receptors. Blood 2012, 120, 4182–4190. [Google Scholar] [CrossRef] [PubMed]
- Corso, J.; Pan, K.T.; Walter, R.; Doebele, C.; Mohr, S.; Bohnenberger, H.; Ströbel, P.; Lenz, C.; Slabicki, M.; Hüllein, J.; et al. Elucidation of tonic and activated B-cell receptor signaling in Burkitt’s lymphoma provides insights into regulation of cell survival. Proc. Natl. Acad. Sci. USA 2016, 113, 5688–5693. [Google Scholar] [CrossRef] [PubMed]
- Noy, A.; de Vos, S.; Thieblemont, C.; Martin, P.; Flowers, C.R.; Morschhauser, F.; Collins, G.P.; Ma, S.; Coleman, M.; Peles, S.; et al. Targeting Bruton tyrosine kinase with ibrutinib in relapsed/refractory marginal zone lymphoma. Blood 2017, 129, 2224–2232. [Google Scholar] [CrossRef] [PubMed]
- Bogusz, A.M.; Baxter, R.H.; Currie, T.; Sinha, P.; Sohani, A.R.; Kutok, J.L.; Rodig, S.J. Quantitative immunofluorescence reveals the signature of active B-cell receptor signaling in diffuse large B-cell lymphoma. Clin. Cancer Res. 2012, 18, 6122–6135. [Google Scholar] [CrossRef]
- Davis, R.E.; Ngo, V.N.; Lenz, G.; Tolar, P.; Young, R.M.; Romesser, P.B.; Kohlhammer, H.; Lamy, L.; Zhao, H.; Yang, Y.; et al. Chronic active B-cell-receptor signalling in diffuse large B-cell lymphoma. Nature 2010, 463, 88–92. [Google Scholar] [CrossRef]
- Phelan, J.D.; Young, R.M.; Webster, D.E.; Roulland, S.; Wright, G.W.; Kasbekar, M.; Shaffer, A.L., 3rd; Ceribelli, M.; Wang, J.Q.; Schmitz, R.; et al. A multiprotein supercomplex controlling oncogenic signalling in lymphoma. Nature 2018, 560, 387–391. [Google Scholar] [CrossRef]
- Havranek, O.; Xu, J.; Köhrer, S.; Wang, Z.; Becker, L.; Comer, J.M.; Henderson, J.; Ma, W.; Man Chun Ma, J.; Westin, J.R.; et al. Tonic B-cell receptor signaling in diffuse large B-cell lymphoma. Blood 2017, 130, 995–1006. [Google Scholar] [CrossRef]
- Myers, D.R.; Zikherman, J.; Roose, J.P. Tonic Signals: Why Do Lymphocytes Bother? Trends Immunol. 2017, 38, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.A.; Eisen, M.B.; Davis, R.E.; Ma, C.; Lossos, I.S.; Rosenwald, A.; Boldrick, J.C.; Sabet, H.; Tran, T.; Yu, X.; et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 2000, 403, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Frustaci, A.M.; Deodato, M.; Zamprogna, G.; Cairoli, R.; Montillo, M.; Tedeschi, A. Next Generation BTK Inhibitors in CLL: Evolving Challenges and New Opportunities. Cancers 2023, 15, 1504. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, L.; Fernández-Miranda, I.; Pérez-Callejo, D.; Quero, C.; Rodríguez, M.; Martín-Acosta, P.; Gómez, S.; González-Rincón, J.; Santos, A.; Tarin, C.; et al. Proposal and validation of a method to classify genetic subtypes of diffuse large B cell lymphoma. Sci. Rep. 2021, 11, 1886. [Google Scholar] [CrossRef] [PubMed]
- Lacy, S.E.; Barrans, S.L.; Beer, P.A.; Painter, D.; Smith, A.G.; Roman, E.; Cooke, S.L.; Ruiz, C.; Glover, P.; Van Hoppe, S.J.L.; et al. Targeted sequencing in DLBCL, molecular subtypes, and outcomes: A Haematological Malignancy Research Network report. Blood 2020, 135, 1759–1771. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.W.; Huang, D.W.; Phelan, J.D.; Coulibaly, Z.A.; Roulland, S.; Young, R.M.; Wang, J.Q.; Schmitz, R.; Morin, R.D.; Tang, J.; et al. A Probabilistic Classification Tool for Genetic Subtypes of Diffuse Large B Cell Lymphoma with Therapeutic Implications. Cancer Cell 2020, 37, 551–568.e14. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.; Wright, G.W.; Huang, D.W.; Johnson, C.A.; Phelan, J.D.; Wang, J.Q.; Roulland, S.; Kasbekar, M.; Young, R.M.; Shaffer, A.L.; et al. Genetics and Pathogenesis of Diffuse Large B-Cell Lymphoma. N. Engl. J. Med. 2018, 378, 1396–1407. [Google Scholar] [CrossRef]
- Chapuy, B.; Stewart, C.; Dunford, A.J.; Kim, J.; Kamburov, A.; Redd, R.A.; Lawrence, M.S.; Roemer, M.G.M.; Li, A.J.; Ziepert, M.; et al. Molecular subtypes of diffuse large B cell lymphoma are associated with distinct pathogenic mechanisms and outcomes. Nat. Med. 2018, 24, 679–690. [Google Scholar] [CrossRef]
- Ma, X.; Zhu, Y.; Dong, D.; Chen, Y.; Wang, S.; Yang, D.; Ma, Z.; Zhang, A.; Zhang, F.; Guo, C.; et al. Cryo-EM structures of two human B cell receptor isotypes. Science 2022, 377, 880–885. [Google Scholar] [CrossRef]
- Su, Q.; Chen, M.; Shi, Y.; Zhang, X.; Huang, G.; Huang, B.; Liu, D.; Liu, Z.; Shi, Y. Cryo-EM structure of the human IgM B cell receptor. Science 2022, 377, 875–880. [Google Scholar] [CrossRef]
- Dong, Y.; Pi, X.; Bartels-Burgahn, F.; Saltukoglu, D.; Liang, Z.; Yang, J.; Alt, F.W.; Reth, M.; Wu, H. Structural principles of B cell antigen receptor assembly. Nature 2022, 612, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Reth, M.; Nitschke, L.; Hikida, M.; Kurosaki, T. Chapter 10—Structure and Signaling Function of the B-Cell Antigen Receptor and Its Coreceptors. In Molecular Biology of B Cells, 2nd ed.; Alt, F.W., Honjo, T., Radbruch, A., Reth, M., Eds.; Academic Press: London, UK, 2015; pp. 151–170. [Google Scholar]
- Tolar, P.; Pierce, S.K. Unveiling the B cell receptor structure. Science 2022, 377, 819–820. [Google Scholar] [CrossRef] [PubMed]
- Tolar, P.; Sohn, H.W.; Pierce, S.K. The initiation of antigen-induced B cell antigen receptor signaling viewed in living cells by fluorescence resonance energy transfer. Nat. Immunol. 2005, 6, 1168–1176. [Google Scholar] [CrossRef] [PubMed]
- Friess, M.D.; Pluhackova, K.; Böckmann, R.A. Structural Model of the mIgM B-Cell Receptor Transmembrane Domain From Self-Association Molecular Dynamics Simulations. Front. Immunol. 2018, 9, 2947. [Google Scholar] [CrossRef] [PubMed]
- Lutz, J.; Dittmann, K.; Bösl, M.R.; Winkler, T.H.; Wienands, J.; Engels, N. Reactivation of IgG-switched memory B cells by BCR-intrinsic signal amplification promotes IgG antibody production. Nat. Commun. 2015, 6, 8575. [Google Scholar] [CrossRef] [PubMed]
- James, L.K. B cells defined by immunoglobulin isotypes. Clin. Exp. Immunol. 2022, 210, 230–239. [Google Scholar] [CrossRef]
- Engels, N.; König, L.M.; Schulze, W.; Radtke, D.; Vanshylla, K.; Lutz, J.; Winkler, T.H.; Nitschke, L.; Wienands, J. The immunoglobulin tail tyrosine motif upgrades memory-type BCRs by incorporating a Grb2-Btk signalling module. Nat. Commun. 2014, 5, 5456. [Google Scholar] [CrossRef]
- Vanshylla, K.; Bartsch, C.; Hitzing, C.; Krümpelmann, L.; Wienands, J.; Engels, N. Grb2 and GRAP connect the B cell antigen receptor to Erk MAP kinase activation in human B cells. Sci. Rep. 2018, 8, 4244. [Google Scholar] [CrossRef]
- Geisberger, R.; Crameri, R.; Achatz, G. Models of signal transduction through the B-cell antigen receptor. Immunology 2003, 110, 401–410. [Google Scholar] [CrossRef]
- Maity, P.C.; Datta, M.; Nicolò, A.; Jumaa, H. Isotype Specific Assembly of B Cell Antigen Receptors and Synergism With Chemokine Receptor CXCR4. Front. Immunol. 2018, 9, 2988. [Google Scholar] [CrossRef]
- Lockey, C.; Young, H.; Brown, J.; Dixon, A.M. Characterization of interactions within the Igα/Igβ transmembrane domains of the human B-cell receptor provides insights into receptor assembly. J. Biol. Chem. 2022, 298, 101843. [Google Scholar] [CrossRef] [PubMed]
- Wemlinger, S.M.; Parker Harp, C.R.; Yu, B.; Hardy, I.R.; Seefeldt, M.; Matsuda, J.; Mingueneau, M.; Spilker, K.A.; Cameron, T.O.; Larrick, J.W.; et al. Preclinical Analysis of Candidate Anti-Human CD79 Therapeutic Antibodies Using a Humanized CD79 Mouse Model. J. Immunol. 2022, 208, 1566–1584. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Wucherpfennig, K.; Patel, D.J. A structural platform for B cell receptor signaling. Cell Res. 2022, 33, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, W.; Ma, J.; Xu, Z. A Novel and Validated 8-Pyroptosis-Related Genes Based Risk Prediction Model for Diffuse Large B Cell Lymphoma. Biomolecules 2022, 12, 1835. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Kläsener, K.; Iype, J.M.; Becker, M.; Maity, P.C.; Cavallari, M.; Nielsen, P.J.; Yang, J.; Reth, M. Continuous signaling of CD79b and CD19 is required for the fitness of Burkitt lymphoma B cells. EMBO J. 2018, 37, e97980. [Google Scholar] [CrossRef] [PubMed]
- Luger, D.; Yang, Y.A.; Raviv, A.; Weinberg, D.; Banerjee, S.; Lee, M.J.; Trepel, J.; Yang, L.; Wakefield, L.M. Expression of the B-cell receptor component CD79a on immature myeloid cells contributes to their tumor promoting effects. PLoS ONE 2013, 8, e76115. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.R. The immunobiology of ubiquitin-dependent B cell receptor functions. Mol. Immunol. 2018, 101, 146–154. [Google Scholar] [CrossRef]
- Corneth, O.B.J.; Neys, S.F.H.; Hendriks, R.W. Aberrant B Cell Signaling in Autoimmune Diseases. Cells 2022, 11, 3391. [Google Scholar] [CrossRef]
- Cyster, J.G.; Allen, C.D.C. B Cell Responses: Cell Interaction Dynamics and Decisions. Cell 2019, 177, 524–540. [Google Scholar] [CrossRef]
- Smith, L.D.; Minton, A.R.; Blunt, M.D.; Karydis, L.I.; Dutton, D.A.; Rogers-Broadway, K.R.; Dobson, R.; Liu, R.; Norster, F.; Hogg, E.; et al. BCR signaling contributes to autophagy regulation in chronic lymphocytic leukemia. Leukemia 2020, 34, 640–644. [Google Scholar] [CrossRef]
- Carter, M.J.; Cox, K.L.; Blakemore, S.J.; Bogdanov, Y.D.; Happo, L.; Scott, C.L.; Strasser, A.; Packham, G.K.; Cragg, M.S. BCR-signaling-induced cell death demonstrates dependency on multiple BH3-only proteins in a murine model of B-cell lymphoma. Cell Death Differ. 2016, 23, 303–312. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, S.K.; Getahun, A.; Gauld, S.B.; Merrell, K.T.; Tamir, I.; Smith, M.J.; Dal Porto, J.M.; Li, Q.Z.; Cambier, J.C. Monophosphorylation of CD79a and CD79b ITAM motifs initiates a SHIP-1 phosphatase-mediated inhibitory signaling cascade required for B cell anergy. Immunity 2011, 35, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Vuillier, F.; Dumas, G.; Magnac, C.; Prevost, M.C.; Lalanne, A.I.; Oppezzo, P.; Melanitou, E.; Dighiero, G.; Payelle-Brogard, B. Lower levels of surface B-cell-receptor expression in chronic lymphocytic leukemia are associated with glycosylation and folding defects of the mu and CD79a chains. Blood 2005, 105, 2933–2940. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Veselits, M.; O’Neill, S.; Hou, P.; Reddi, A.L.; Berlin, I.; Ikeda, M.; Nash, P.D.; Longnecker, R.; Band, H.; et al. Ubiquitinylation of Ig beta dictates the endocytic fate of the B cell antigen receptor. J. Immunol. 2007, 179, 4435–4443. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Iwai, K. Roles of the NF-κB Pathway in B-Lymphocyte Biology. Curr. Top. Microbiol. Immunol. 2016, 393, 177–209. [Google Scholar] [CrossRef] [PubMed]
- Almaden, J.V.; Liu, Y.C.; Yang, E.; Otero, D.C.; Birnbaum, H.; Davis-Turak, J.; Asagiri, M.; David, M.; Goldrath, A.W.; Hoffmann, A. B-cell survival and development controlled by the coordination of NF-κB family members RelB and cRel. Blood 2016, 127, 1276–1286. [Google Scholar] [CrossRef]
- Szydłowski, M.; Jabłońska, E.; Juszczyński, P. FOXO1 transcription factor: A critical effector of the PI3K-AKT axis in B-cell development. Int. Rev. Immunol. 2014, 33, 146–157. [Google Scholar] [CrossRef]
- Chen, J.; Limon, J.J.; Blanc, C.; Peng, S.L.; Fruman, D.A. Foxo1 regulates marginal zone B-cell development. Eur. J. Immunol. 2010, 40, 1890–1896. [Google Scholar] [CrossRef]
- Sevdali, E.; Block, V.; Lataretu, M.; Li, H.; Smulski, C.R.; Briem, J.S.; Heitz, Y.; Fischer, B.; Ramirez, N.J.; Grimbacher, B.; et al. BAFFR activates PI3K/AKT signaling in human naive but not in switched memory B cells through direct interactions with B cell antigen receptors. Cell Rep. 2022, 39, 111019. [Google Scholar] [CrossRef]
- Lang, P.; Stolpa, J.C.; Freiberg, B.A.; Crawford, F.; Kappler, J.; Kupfer, A.; Cambier, J.C. TCR-induced transmembrane signaling by peptide/MHC class II via associated Ig-alpha/beta dimers. Science 2001, 291, 1537–1540. [Google Scholar] [CrossRef]
- Katikaneni, D.S.; Jin, L. B cell MHC class II signaling: A story of life and death. Hum. Immunol. 2019, 80, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Ichinose, S.; Hayashizaki, K.; Tsubata, T. Induction of autophagy by B cell antigen receptor stimulation and its inhibition by costimulation. Biochem. Biophys. Res. Commun. 2008, 374, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Arbogast, F.; Arnold, J.; Hammann, P.; Kuhn, L.; Chicher, J.; Murera, D.; Weishaar, J.; Muller, S.; Fauny, J.D.; Gros, F. ATG5 is required for B cell polarization and presentation of particulate antigens. Autophagy 2019, 15, 280–294. [Google Scholar] [CrossRef] [PubMed]
- Caro-Maldonado, A.; Wang, R.; Nichols, A.G.; Kuraoka, M.; Milasta, S.; Sun, L.D.; Gavin, A.L.; Abel, E.D.; Kelsoe, G.; Green, D.R.; et al. Metabolic reprogramming is required for antibody production that is suppressed in anergic but exaggerated in chronically BAFF-exposed B cells. J. Immunol. 2014, 192, 3626–3636. [Google Scholar] [CrossRef] [PubMed]
- Jumaa, H.; Caganova, M.; McAllister, E.J.; Hoenig, L.; He, X.; Saltukoglu, D.; Brenker, K.; Köhler, M.; Leben, R.; Hauser, A.E.; et al. Immunoglobulin expression in the endoplasmic reticulum shapes the metabolic fitness of B lymphocytes. Life Sci. Alliance 2020, 3, e202000700. [Google Scholar] [CrossRef] [PubMed]
- Love, P.E.; Hayes, S.M. ITAM-mediated signaling by the T-cell antigen receptor. Cold Spring Harb. Perspect. Biol. 2010, 2, a002485. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.W., Jr.; Imboden, J.B.; Torres, R.M. Antigen Receptor Genes, Gene Products, and Coreceptors. In Clinical Immunology: Principles and Practice, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Xu, W.; Berning, P.; Lenz, G. Targeting B-cell receptor and PI3K signaling in diffuse large B-cell lymphoma. Blood 2021, 138, 1110–1119. [Google Scholar] [CrossRef]
- Roche, P.A.; Furuta, K. The ins and outs of MHC class II-mediated antigen processing and presentation. Nat. Rev. Immunol. 2015, 15, 203–216. [Google Scholar] [CrossRef]
- Busman-Sahay, K.; Drake, L.; Sitaram, A.; Marks, M.; Drake, J.R. Cis and trans regulatory mechanisms control AP2-mediated B cell receptor endocytosis via select tyrosine-based motifs. PLoS ONE 2013, 8, e54938. [Google Scholar] [CrossRef]
- Crute, B.W.; Sheraden, R.; Ott, V.L.; Harley, I.T.W.; Getahun, A.; Cambier, J.C. Inhibitory Receptor Trap: A Platform for Discovery of Inhibitory Receptors That Utilize Inositol Lipid and Phosphotyrosine Phosphatase Effectors. Front. Immunol. 2020, 11, 592329. [Google Scholar] [CrossRef]
- Franks, S.E.; Cambier, J.C. Putting on the Brakes: Regulatory Kinases and Phosphatases Maintaining B Cell Anergy. Front. Immunol. 2018, 9, 665. [Google Scholar] [CrossRef] [PubMed]
- Pao, L.I.; Famiglietti, S.J.; Cambier, J.C. Asymmetrical phosphorylation and function of immunoreceptor tyrosine-based activation motif tyrosines in B cell antigen receptor signal transduction. J. Immunol. 1998, 160, 3305–3314. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.R.; Campbell, K.S.; Kazlauskas, A.; Johnson, S.A.; Hertz, M.; Potter, T.A.; Pleiman, C.; Cambier, J.C. The B cell antigen receptor complex: Association of Ig-alpha and Ig-beta with distinct cytoplasmic effectors. Science 1992, 258, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Mkaddem, S.B.; Murua, A.; Flament, H.; Titeca-Beauport, D.; Bounaix, C.; Danelli, L.; Launay, P.; Benhamou, M.; Blank, U.; Daugas, E.; et al. Lyn and Fyn function as molecular switches that control immunoreceptors to direct homeostasis or inflammation. Nat. Commun. 2017, 8, 246. [Google Scholar] [CrossRef] [PubMed]
- Gross, A.J.; Lyandres, J.R.; Panigrahi, A.K.; Prak, E.T.; DeFranco, A.L. Developmental acquisition of the Lyn-CD22-SHP-1 inhibitory pathway promotes B cell tolerance. J. Immunol. 2009, 182, 5382–5392. [Google Scholar] [CrossRef]
- Khan, N.; Rothstein, T.L. The Alternate Pathway for BCR Signaling Induced by IL-4 Requires Lyn Tyrosine Kinase. J. Mol. Biol. 2021, 433, 166667. [Google Scholar] [CrossRef]
- Kohlhas, V.; Hallek, M.; Nguyen, P.H. Constitutive activation of Lyn kinase enhances BCR responsiveness, but not the development of CLL in Eµ-TCL1 mice. Blood Adv. 2020, 4, 6106–6116. [Google Scholar] [CrossRef]
- Alsadeq, A.; Hobeika, E.; Medgyesi, D.; Kläsener, K.; Reth, M. The role of the Syk/Shp-1 kinase-phosphatase equilibrium in B cell development and signaling. J. Immunol. 2014, 193, 268–276. [Google Scholar] [CrossRef]
- Adachi, T.; Wienands, J.; Wakabayashi, C.; Yakura, H.; Reth, M.; Tsubata, T. SHP-1 requires inhibitory co-receptors to down-modulate B cell antigen receptor-mediated phosphorylation of cellular substrates. J. Biol. Chem. 2001, 276, 26648–26655. [Google Scholar] [CrossRef]
- Miyazaki, A.; Yogosawa, S.; Murakami, A.; Kitamura, D. Identification of CMTM7 as a transmembrane linker of BLNK and the B-cell receptor. PLoS ONE 2012, 7, e31829. [Google Scholar] [CrossRef]
- Patterson, H.C.; Kraus, M.; Kim, Y.M.; Ploegh, H.; Rajewsky, K. The B cell receptor promotes B cell activation and proliferation through a non-ITAM tyrosine in the Igalpha cytoplasmic domain. Immunity 2006, 25, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Kabak, S.; Skaggs, B.J.; Gold, M.R.; Affolter, M.; West, K.L.; Foster, M.S.; Siemasko, K.; Chan, A.C.; Aebersold, R.; Clark, M.R. The direct recruitment of BLNK to immunoglobulin alpha couples the B-cell antigen receptor to distal signaling pathways. Mol. Cell Biol. 2002, 22, 2524–2535. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Phelan, J.D.; Wright, G.W.; Häupl, B.; Huang, D.W.; Shaffer, A.L., 3rd; Young, R.M.; Wang, Z.; Zhao, H.; Yu, X.; et al. Regulation of B cell receptor-dependent NF-κB signaling by the tumor suppressor KLHL14. Proc. Natl. Acad. Sci. USA 2020, 117, 6092–6102. [Google Scholar] [CrossRef] [PubMed]
- Veselits, M.; Tanaka, A.; Chen, Y.; Hamel, K.; Mandal, M.; Kandasamy, M.; Manicassamy, B.; O’Neill, S.K.; Wilson, P.; Sciammas, R.; et al. Igβ ubiquitination activates PI3K signals required for endosomal sorting. J. Exp. Med. 2017, 214, 3775–3790. [Google Scholar] [CrossRef] [PubMed]
- Satpathy, S.; Wagner, S.A.; Beli, P.; Gupta, R.; Kristiansen, T.A.; Malinova, D.; Francavilla, C.; Tolar, P.; Bishop, G.A.; Hostager, B.S.; et al. Systems-wide analysis of BCR signalosomes and downstream phosphorylation and ubiquitylation. Mol. Syst. Biol. 2015, 11, 810. [Google Scholar] [CrossRef] [PubMed]
- Drake, L.; McGovern-Brindisi, E.M.; Drake, J.R. BCR ubiquitination controls BCR-mediated antigen processing and presentation. Blood 2006, 108, 4086–4093. [Google Scholar] [CrossRef] [PubMed]
- Katkere, B.; Rosa, S.; Drake, J.R. The Syk-binding ubiquitin ligase c-Cbl mediates signaling-dependent B cell receptor ubiquitination and B cell receptor-mediated antigen processing and presentation. J. Biol. Chem. 2012, 287, 16636–16644. [Google Scholar] [CrossRef]
- Fearon, D.T.; Carroll, M.C. Regulation of B lymphocyte responses to foreign and self-antigens by the CD19/CD21 complex. Annu. Rev. Immunol. 2000, 18, 393–422. [Google Scholar] [CrossRef]
- Whillock, A.L.; Ybarra, T.K.; Bishop, G.A. TNF receptor-associated factor 3 restrains B-cell receptor signaling in normal and malignant B cells. J. Biol. Chem. 2021, 296, 100465. [Google Scholar] [CrossRef]
- Bishop, G.A.; Stunz, L.L.; Hostager, B.S. TRAF3 as a Multifaceted Regulator of B Lymphocyte Survival and Activation. Front. Immunol. 2018, 9, 2161. [Google Scholar] [CrossRef]
- Vallabhapurapu, S.; Matsuzawa, A.; Zhang, W.; Tseng, P.H.; Keats, J.J.; Wang, H.; Vignali, D.A.; Bergsagel, P.L.; Karin, M. Nonredundant and complementary functions of TRAF2 and TRAF3 in a ubiquitination cascade that activates NIK-dependent alternative NF-kappaB signaling. Nat. Immunol. 2008, 9, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Mosquera Orgueira, A.; Ferreiro Ferro, R.; Díaz Arias, J.; Aliste Santos, C.; Antelo Rodríguez, B.; Bao Pérez, L.; Alonso Vence, N.; Bendaña López, Á.; Abuin Blanco, A.; Melero Valentín, P.; et al. Detection of new drivers of frequent B-cell lymphoid neoplasms using an integrated analysis of whole genomes. PLoS ONE 2021, 16, e0248886. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.H.; Young, R.M.; Schmitz, R.; Yang, Y.; Pittaluga, S.; Wright, G.; Lih, C.J.; Williams, P.M.; Shaffer, A.L.; Gerecitano, J.; et al. Targeting B cell receptor signaling with ibrutinib in diffuse large B cell lymphoma. Nat. Med. 2015, 21, 922–926. [Google Scholar] [CrossRef] [PubMed]
- Visco, C.; Tanasi, I.; Quaglia, F.M.; Ferrarini, I.; Fraenza, C.; Krampera, M. Oncogenic Mutations of MYD88 and CD79B in Diffuse Large B-Cell Lymphoma and Implications for Clinical Practice. Cancers 2020, 12, 2913. [Google Scholar] [CrossRef] [PubMed]
- Grondona, P.; Bucher, P.; Schulze-Osthoff, K.; Hailfinger, S.; Schmitt, A. NF-κB Activation in Lymphoid Malignancies: Genetics, Signaling, and Targeted Therapy. Biomedicines 2018, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Miao, Y.; Medeiros, L.J.; Xu-Monette, Z.Y.; Li, J.; Young, K.H. Dysregulation of Cell Survival in Diffuse Large B Cell Lymphoma: Mechanisms and Therapeutic Targets. Front. Oncol. 2019, 9, 107. [Google Scholar] [CrossRef] [PubMed]
- Nagel, D.; Bognar, M.; Eitelhuber, A.C.; Kutzner, K.; Vincendeau, M.; Krappmann, D. Combinatorial BTK and MALT1 inhibition augments killing of CD79 mutant diffuse large B cell lymphoma. Oncotarget 2015, 6, 42232–42242. [Google Scholar] [CrossRef]
- Pasqualucci, L.; Klein, U. NF-κB Mutations in Germinal Center B-Cell Lymphomas: Relation to NF-κB Function in Normal B Cells. Biomedicines 2022, 10, 2450. [Google Scholar] [CrossRef]
- Ducharme, O.; Beylot-Barry, M.; Pham-Ledard, A.; Bohers, E.; Viailly, P.J.; Bandres, T.; Faur, N.; Frison, E.; Vergier, B.; Jardin, F.; et al. Mutations of the B-Cell Receptor Pathway Confer Chemoresistance in Primary Cutaneous Diffuse Large B-Cell Lymphoma Leg Type. J. Investig. Dermatol. 2019, 139, 2334–2342.e8. [Google Scholar] [CrossRef]
- Myklebust, J.H.; Brody, J.; Kohrt, H.E.; Kolstad, A.; Czerwinski, D.K.; Wälchli, S.; Green, M.R.; Trøen, G.; Liestøl, K.; Beiske, K.; et al. Distinct patterns of B-cell receptor signaling in non-Hodgkin lymphomas identified by single-cell profiling. Blood 2017, 129, 759–770. [Google Scholar] [CrossRef]
- Thurner, L.; Hartmann, S.; Neumann, F.; Hoth, M.; Stilgenbauer, S.; Küppers, R.; Preuss, K.D.; Bewarder, M. Role of Specific B-Cell Receptor Antigens in Lymphomagenesis. Front. Oncol. 2020, 10, 604685. [Google Scholar] [CrossRef] [PubMed]
- Andrades, A.; Álvarez-Pérez, J.C.; Patiño-Mercau, J.R.; Cuadros, M.; Baliñas-Gavira, C.; Medina, P.P. Recurrent splice site mutations affect key diffuse large B-cell lymphoma genes. Blood 2022, 139, 2406–2410. [Google Scholar] [CrossRef] [PubMed]
- Young, R.M.; Wu, T.; Schmitz, R.; Dawood, M.; Xiao, W.; Phelan, J.D.; Xu, W.; Menard, L.; Meffre, E.; Chan, W.C.; et al. Survival of human lymphoma cells requires B-cell receptor engagement by self-antigens. Proc. Natl. Acad. Sci. USA 2015, 112, 13447–13454. [Google Scholar] [CrossRef] [PubMed]
- Lohr, J.G.; Stojanov, P.; Lawrence, M.S.; Auclair, D.; Chapuy, B.; Sougnez, C.; Cruz-Gordillo, P.; Knoechel, B.; Asmann, Y.W.; Slager, S.L.; et al. Discovery and prioritization of somatic mutations in diffuse large B-cell lymphoma (DLBCL) by whole-exome sequencing. Proc. Natl. Acad. Sci. USA 2012, 109, 3879–3884. [Google Scholar] [CrossRef] [PubMed]
- Payelle-Brogard, B.; Magnac, C.; Mauro, F.R.; Mandelli, F.; Dighiero, G. Analysis of the B-cell receptor B29 (CD79b) gene in familial chronic lymphocytic leukemia. Blood 1999, 94, 3516–3522. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Yamaguchi, M.; Kobayashi, K.; Miyazaki, K.; Tawara, I.; Imai, H.; Ono, R.; Nosaka, T.; Tanaka, K.; Katayama, N. MYD88, CD79B, and CARD11 gene mutations in CD5-positive diffuse large B-cell lymphoma. Cancer 2017, 123, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Bohers, E.; Mareschal, S.; Bouzelfen, A.; Marchand, V.; Ruminy, P.; Maingonnat, C.; Ménard, A.L.; Etancelin, P.; Bertrand, P.; Dubois, S.; et al. Targetable activating mutations are very frequent in GCB and ABC diffuse large B-cell lymphoma. Genes Chromosomes Cancer 2014, 53, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Poulain, S.; Roumier, C.; Galiègue-Zouitina, S.; Daudignon, A.; Herbaux, C.; Aiijou, R.; Lainelle, A.; Broucqsault, N.; Bertrand, E.; Manier, S.; et al. Genome wide SNP array identified multiple mechanisms of genetic changes in Waldenstrom macroglobulinemia. Am. J. Hematol. 2013, 88, 948–954. [Google Scholar] [CrossRef]
- Okosun, J.; Bödör, C.; Wang, J.; Araf, S.; Yang, C.Y.; Pan, C.; Boller, S.; Cittaro, D.; Bozek, M.; Iqbal, S.; et al. Integrated genomic analysis identifies recurrent mutations and evolution patterns driving the initiation and progression of follicular lymphoma. Nat. Genet. 2014, 46, 176–181. [Google Scholar] [CrossRef]
- Krysiak, K.; Gomez, F.; White, B.S.; Matlock, M.; Miller, C.A.; Trani, L.; Fronick, C.C.; Fulton, R.S.; Kreisel, F.; Cashen, A.F.; et al. Recurrent somatic mutations affecting B-cell receptor signaling pathway genes in follicular lymphoma. Blood 2017, 129, 473–483. [Google Scholar] [CrossRef]
- Trøen, G.; Warsame, A.; Delabie, J. CD79B and MYD88 Mutations in Splenic Marginal Zone Lymphoma. ISRN Oncol. 2013, 2013, 252318. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.A.; Talley, J.A.; Do, H.N.; Kagan, H.L.; Kunkel, L.; Berenson, J.; Cooper, M.D.; Saxon, A.; Wall, R. Aberrations of the B-Cell Receptor B29 (CD79b) Gene in Chronic Lymphocytic Leukemia. Blood 1997, 90, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Cetin, G.O.; Baris, I.C.; Caner, V.; Sarikepe, B.; Sen Turk, N.; Tepeli, E.; Hacioglu, S.; Sari, I.; Bagci, G.; Keskin, A. Mutational status of EZH2 and CD79B hot spots in mature B-cell non-Hodgkin’s lymphomas: Novel CD79B variations have been revealed. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 830–836. [Google Scholar] [PubMed]
- Bruno, A.; Boisselier, B.; Labreche, K.; Marie, Y.; Polivka, M.; Jouvet, A.; Adam, C.; Figarella-Branger, D.; Miquel, C.; Eimer, S.; et al. Mutational analysis of primary central nervous system lymphoma. Oncotarget 2014, 5, 5065–5075. [Google Scholar] [CrossRef]
- Kim, Y.; Ju, H.; Kim, D.H.; Yoo, H.Y.; Kim, S.J.; Kim, W.S.; Ko, Y.H. CD79B and MYD88 mutations in diffuse large B-cell lymphoma. Hum. Pathol. 2014, 45, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Saieg, M.A.; Geddie, W.R.; Boerner, S.L.; Bailey, D.; Crump, M.; da Cunha Santos, G. EZH2 and CD79B mutational status over time in B-cell non-Hodgkin lymphomas detected by high-throughput sequencing using minimal samples. Cancer Cytopathol. 2013, 121, 377–386. [Google Scholar] [CrossRef]
- Hunter, Z.R.; Xu, L.; Yang, G.; Zhou, Y.; Liu, X.; Cao, Y.; Manning, R.J.; Tripsas, C.; Patterson, C.J.; Sheehy, P.; et al. The genomic landscape of Waldenström macroglobulinemia is characterized by highly recurring MYD88 and WHIM-like CXCR4 mutations, and small somatic deletions associated with B-cell lymphomagenesis. Blood 2014, 123, 1637–1646. [Google Scholar] [CrossRef]
- Alfarano, A.; Indraccolo, S.; Circosta, P.; Minuzzo, S.; Vallario, A.; Zamarchi, R.; Fregonese, A.; Calderazzo, F.; Faldella, A.; Aragno, M.; et al. An alternatively spliced form of CD79b gene may account for altered B-cell receptor expression in B-chronic lymphocytic leukemia. Blood 1999, 93, 2327–2335. [Google Scholar] [CrossRef]
- Jiménez, C.; Alonso-Álvarez, S.; Alcoceba, M.; Ordóñez, G.R.; García-Álvarez, M.; Prieto-Conde, M.I.; Chillón, M.C.; Balanzategui, A.; Corral, R.; Marín, L.A.; et al. From Waldenström’s macroglobulinemia to aggressive diffuse large B-cell lymphoma: A whole-exome analysis of abnormalities leading to transformation. Blood Cancer J. 2017, 7, e591. [Google Scholar] [CrossRef]
- Wang, J.Q.; Jeelall, Y.S.; Humburg, P.; Batchelor, E.L.; Kaya, S.M.; Yoo, H.M.; Goodnow, C.C.; Horikawa, K. Synergistic cooperation and crosstalk between MYD88(L265P) and mutations that dysregulate CD79B and surface IgM. J. Exp. Med. 2017, 214, 2759–2776. [Google Scholar] [CrossRef]
- Gordon, M.S.; Kato, R.M.; Lansigan, F.; Thompson, A.A.; Wall, R.; Rawlings, D.J. Aberrant B cell receptor signaling from B29 (Igbeta, CD79b) gene mutations of chronic lymphocytic leukemia B cells. Proc. Natl. Acad. Sci. USA 2000, 97, 5504–5509. [Google Scholar] [CrossRef] [PubMed]
- Faumont, N.; Taoui, O.; Collares, D.; Jais, J.P.; Leroy, K.; Prévaud, L.; Jardin, F.; Molina, T.J.; Copie-Bergman, C.; Petit, B.; et al. c-Rel Is the Pivotal NF-κB Subunit in Germinal Center Diffuse Large B-Cell Lymphoma: A LYSA Study. Front. Oncol. 2021, 11, 638897. [Google Scholar] [CrossRef] [PubMed]
- Compagno, M.; Lim, W.K.; Grunn, A.; Nandula, S.V.; Brahmachary, M.; Shen, Q.; Bertoni, F.; Ponzoni, M.; Scandurra, M.; Califano, A.; et al. Mutations of multiple genes cause deregulation of NF-kappaB in diffuse large B-cell lymphoma. Nature 2009, 459, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.E.; Brown, K.D.; Siebenlist, U.; Staudt, L.M. Constitutive nuclear factor kappaB activity is required for survival of activated B cell-like diffuse large B cell lymphoma cells. J. Exp. Med. 2001, 194, 1861–1874. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.Y.; Bauman, B.M.; Arjunaraja, S.; Dorjbal, B.; Milner, J.D.; Snow, A.L.; Turvey, S.E. The CBM-opathies-A Rapidly Expanding Spectrum of Human Inborn Errors of Immunity Caused by Mutations in the CARD11-BCL10-MALT1 Complex. Front. Immunol. 2018, 9, 2078. [Google Scholar] [CrossRef] [PubMed]
- Turvey, S.E.; Durandy, A.; Fischer, A.; Fung, S.Y.; Geha, R.S.; Gewies, A.; Giese, T.; Greil, J.; Keller, B.; McKinnon, M.L.; et al. The CARD11-BCL10-MALT1 (CBM) signalosome complex: Stepping into the limelight of human primary immunodeficiency. J. Allergy Clin. Immunol. 2014, 134, 276–284. [Google Scholar] [CrossRef]
- Kloo, B.; Nagel, D.; Pfeifer, M.; Grau, M.; Düwel, M.; Vincendeau, M.; Dörken, B.; Lenz, P.; Lenz, G.; Krappmann, D. Critical role of PI3K signaling for NF-kappaB-dependent survival in a subset of activated B-cell-like diffuse large B-cell lymphoma cells. Proc. Natl. Acad. Sci. USA 2011, 108, 272–277. [Google Scholar] [CrossRef]
- Kim, A.; Seong, K.M.; Kang, H.J.; Park, S.; Lee, S.S. Inhibition of Lyn is a promising treatment for mantle cell lymphoma with bortezomib resistance. Oncotarget 2015, 6, 38225–38238. [Google Scholar] [CrossRef]
- Ezell, S.A.; Wang, S.; Bihani, T.; Lai, Z.; Grosskurth, S.E.; Tepsuporn, S.; Davies, B.R.; Huszar, D.; Byth, K.F. Differential regulation of mTOR signaling determines sensitivity to AKT inhibition in diffuse large B cell lymphoma. Oncotarget 2016, 7, 9163–9174. [Google Scholar] [CrossRef]
- Majchrzak, A.; Witkowska, M.; Smolewski, P. Inhibition of the PI3K/Akt/mTOR signaling pathway in diffuse large B-cell lymphoma: Current knowledge and clinical significance. Molecules 2014, 19, 14304–14315. [Google Scholar] [CrossRef]
- Ma, M.C.J.; Tadros, S.; Bouska, A.; Heavican, T.; Yang, H.; Deng, Q.; Moore, D.; Akhter, A.; Hartert, K.; Jain, N.; et al. Subtype-specific and co-occurring genetic alterations in B-cell non-Hodgkin lymphoma. Haematologica 2022, 107, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.P.; Shen, R.; Shi, Z.Y.; Cheng, S.; Wang, L.; Liu, Y.; Zhang, L.; Huang, R.; Ma, X.; Wu, X.; et al. The Prognostic Significance of CD79B Mutation in Diffuse Large B-Cell Lymphoma: A Meta-analysis and Systematic Literature Review. Clin. Lymphoma Myeloma Leuk. 2022, 22, e1051–e1058.e1. [Google Scholar] [CrossRef] [PubMed]
- de Groen, R.A.L.; Schrader, A.M.R.; Kersten, M.J.; Pals, S.T.; Vermaat, J.S.P. MYD88 in the driver’s seat of B-cell lymphomagenesis: From molecular mechanisms to clinical implications. Haematologica 2019, 104, 2337–2348. [Google Scholar] [CrossRef] [PubMed]
- Sewastianik, T.; Guerrera, M.L.; Adler, K.; Dennis, P.S.; Wright, K.; Shanmugam, V.; Huang, Y.; Tanton, H.; Jiang, M.; Kofides, A.; et al. Human MYD88L265P is insufficient by itself to drive neoplastic transformation in mature mouse B cells. Blood Adv. 2019, 3, 3360–3374. [Google Scholar] [CrossRef] [PubMed]
- Alcoceba, M.; García-Álvarez, M.; Medina, A.; Maldonado, R.; González-Calle, V.; Chillón, M.C.; Sarasquete, M.E.; González, M.; García-Sanz, R.; Jiménez, C. MYD88 Mutations: Transforming the Landscape of IgM Monoclonal Gammopathies. Int. J. Mol. Sci. 2022, 23, 5570. [Google Scholar] [CrossRef] [PubMed]
- Motshwene, P.G.; Moncrieffe, M.C.; Grossmann, J.G.; Kao, C.; Ayaluru, M.; Sandercock, A.M.; Robinson, C.V.; Latz, E.; Gay, N.J. An oligomeric signaling platform formed by the Toll-like receptor signal transducers MyD88 and IRAK-4. J. Biol. Chem. 2009, 284, 25404–25411. [Google Scholar] [CrossRef] [PubMed]
- Balka, K.R.; De Nardo, D. Understanding early TLR signaling through the Myddosome. J. Leukoc. Biol. 2019, 105, 339–351. [Google Scholar] [CrossRef]
- Cao, F.; Deliz-Aguirre, R.; Gerpott, F.H.; Ziska, E.; Taylor, M.J. Myddosome clustering in IL-1 receptor signaling regulates the formation of an NF-κB activating signalosome. EMBO Rep. 2023, 24, e57233. [Google Scholar] [CrossRef]
- De Nardo, D.; Balka, K.R.; Cardona Gloria, Y.; Rao, V.R.; Latz, E.; Masters, S.L. Interleukin-1 receptor-associated kinase 4 (IRAK4) plays a dual role in myddosome formation and Toll-like receptor signaling. J. Biol. Chem. 2018, 293, 15195–15207. [Google Scholar] [CrossRef]
- Weber, T.; Schmitz, R. Molecular Subgroups of Diffuse Large B Cell Lymphoma: Biology and Implications for Clinical Practice. Curr. Oncol. Rep. 2022, 24, 13–21. [Google Scholar] [CrossRef]
- Roschewski, M.; Phelan, J.D.; Wilson, W.H. Molecular Classification and Treatment of Diffuse Large B-Cell Lymphoma and Primary Mediastinal B-Cell Lymphoma. Cancer J. 2020, 26, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Fornecker, L.M.; Muller, L.; Bertrand, F.; Paul, N.; Pichot, A.; Herbrecht, R.; Chenard, M.P.; Mauvieux, L.; Vallat, L.; Bahram, S.; et al. Multi-omics dataset to decipher the complexity of drug resistance in diffuse large B-cell lymphoma. Sci. Rep. 2019, 9, 895. [Google Scholar] [CrossRef] [PubMed]
- George, B.; Chowdhury, S.M.; Hart, A.; Sircar, A.; Singh, S.K.; Nath, U.K.; Mamgain, M.; Singhal, N.K.; Sehgal, L.; Jain, N. Ibrutinib Resistance Mechanisms and Treatment Strategies for B-Cell lymphomas. Cancers 2020, 12, 1328. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, W.S.; Ryu, K.; Kim, S.J.; Park, C. CD79B limits response of diffuse large B cell lymphoma to ibrutinib. Leuk. Lymphoma 2016, 57, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Rip, J.; de Bruijn, M.J.W.; Neys, S.F.H.; Singh, S.P.; Willar, J.; van Hulst, J.A.C.; Hendriks, R.W.; Corneth, O.B.J. Bruton’s tyrosine kinase inhibition induces rewiring of proximal and distal B-cell receptor signaling in mice. Eur. J. Immunol. 2021, 51, 2251–2265. [Google Scholar] [CrossRef]
- Domka, K.; Goral, A.; Firczuk, M. cROSsing the Line: Between Beneficial and Harmful Effects of Reactive Oxygen Species in B-Cell Malignancies. Front. Immunol. 2020, 11, 1538. [Google Scholar] [CrossRef]
- Wang, S.S.; Davis, S.; Cerhan, J.R.; Hartge, P.; Severson, R.K.; Cozen, W.; Lan, Q.; Welch, R.; Chanock, S.J.; Rothman, N. Polymorphisms in oxidative stress genes and risk for non-Hodgkin lymphoma. Carcinogenesis 2006, 27, 1828–1834. [Google Scholar] [CrossRef]
- Müller-Winkler, J.; Mitter, R.; Rappe, J.C.F.; Vanes, L.; Schweighoffer, E.; Mohammadi, H.; Wack, A.; Tybulewicz, V.L.J. Critical requirement for BCR, BAFF, and BAFFR in memory B cell survival. J. Exp. Med. 2021, 218, e20191393. [Google Scholar] [CrossRef]
Figure 1.
Distinct function of CD79a/CD79b domains in B-cell receptor signaling. Abbreviations: BCR—B-cell receptor; Fc region—fragment crystallizable region; IgH—immunoglobulin heavy chain; ITAM—immunoreceptor tyrosine-based activation motif. This figure was created with biorender.com.
Figure 1.
Distinct function of CD79a/CD79b domains in B-cell receptor signaling. Abbreviations: BCR—B-cell receptor; Fc region—fragment crystallizable region; IgH—immunoglobulin heavy chain; ITAM—immunoreceptor tyrosine-based activation motif. This figure was created with biorender.com.

Figure 2.
Phosphorylation of ITAM and non-ITAM tyrosines of CD79a and CD79b differentially modulates BCR signaling. Abbreviations: BLNK—B-cell linker protein; BCR—B-cell receptor; BTK—Bruton’s tyrosine kinase; Dok-1—docking protein 1; ERK—extracellular signal-regulated kinase; Grb2—growth factor receptor-bound protein 2; ITAM—immunoreceptor tyrosine-based activation motif; ITIM—immunoreceptor tyrosine-based inhibitory motif; JNK—c-Jun N-terminal kinase; Lyn—Lck/Yes novel tyrosine kinase; NF-κB—nuclear factor kappa-light-chain-enhancer of activated B cells; P—phosphate group; PI3K—phosphoinositide 3-kinase; PLC-γ2—phosphoinositide phospholipase C-γ-2; PTEN—phosphatase and tensin homolog; SHIP-1—Src homology 2 (SH2) domain-containing inositol-5-phosphatase 1; SHP-1—Src homology region 2 domain-containing phosphatase-1; SYK—spleen-associated tyrosine kinase. This figure was created with biorender.com.
Figure 2.
Phosphorylation of ITAM and non-ITAM tyrosines of CD79a and CD79b differentially modulates BCR signaling. Abbreviations: BLNK—B-cell linker protein; BCR—B-cell receptor; BTK—Bruton’s tyrosine kinase; Dok-1—docking protein 1; ERK—extracellular signal-regulated kinase; Grb2—growth factor receptor-bound protein 2; ITAM—immunoreceptor tyrosine-based activation motif; ITIM—immunoreceptor tyrosine-based inhibitory motif; JNK—c-Jun N-terminal kinase; Lyn—Lck/Yes novel tyrosine kinase; NF-κB—nuclear factor kappa-light-chain-enhancer of activated B cells; P—phosphate group; PI3K—phosphoinositide 3-kinase; PLC-γ2—phosphoinositide phospholipase C-γ-2; PTEN—phosphatase and tensin homolog; SHIP-1—Src homology 2 (SH2) domain-containing inositol-5-phosphatase 1; SHP-1—Src homology region 2 domain-containing phosphatase-1; SYK—spleen-associated tyrosine kinase. This figure was created with biorender.com.

Figure 3.
The role of CD79a/CD79b in B-cell receptor signaling in normal B cells. Abbreviations: BAFF—B-cell activating factor; BAFFR—B-cell activating factor receptor; BCR—B-cell receptor; BTK—Bruton’s tyrosine kinase; ERK—extracellular signal-regulated kinase; Grb2—growth factor receptor-bound protein 2; ITAM—immunoreceptor tyrosine-based activation motif; ITIM—immunoreceptor tyrosine-based inhibitory motif; KLHL14—Kelch Like Family Member 14; Lyn—Lck/Yes novel tyrosine kinase; MAPK—mitogen-activated protein kinase; MHC II—major histocompatibility complex class II; NF-κB—nuclear factor kappa-light-chain-enhancer of activated B cells; P—phosphate group; PI3K—phosphoinositide 3-kinase; PIP3—phosphatidylinositol (3,4,5)-trisphosphate; PLCγ2—phosphoinositide phospholipase C-γ-2; PTEN—phosphatase and tensin homolog; SHIP-1—Src homology region 2 domain-containing inositol 5′ phosphatase 1; SHP-1—Src homology region 2 domain-containing tyrosine phosphatase; Syk—spleen-associated tyrosine kinase; TLR9—toll-like receptor 9; TRAF3—tumor necrosis factor receptor (TNFR)-associated factor 3; Ub—ubiquitination. This figure was created with biorender.com.
Figure 3.
The role of CD79a/CD79b in B-cell receptor signaling in normal B cells. Abbreviations: BAFF—B-cell activating factor; BAFFR—B-cell activating factor receptor; BCR—B-cell receptor; BTK—Bruton’s tyrosine kinase; ERK—extracellular signal-regulated kinase; Grb2—growth factor receptor-bound protein 2; ITAM—immunoreceptor tyrosine-based activation motif; ITIM—immunoreceptor tyrosine-based inhibitory motif; KLHL14—Kelch Like Family Member 14; Lyn—Lck/Yes novel tyrosine kinase; MAPK—mitogen-activated protein kinase; MHC II—major histocompatibility complex class II; NF-κB—nuclear factor kappa-light-chain-enhancer of activated B cells; P—phosphate group; PI3K—phosphoinositide 3-kinase; PIP3—phosphatidylinositol (3,4,5)-trisphosphate; PLCγ2—phosphoinositide phospholipase C-γ-2; PTEN—phosphatase and tensin homolog; SHIP-1—Src homology region 2 domain-containing inositol 5′ phosphatase 1; SHP-1—Src homology region 2 domain-containing tyrosine phosphatase; Syk—spleen-associated tyrosine kinase; TLR9—toll-like receptor 9; TRAF3—tumor necrosis factor receptor (TNFR)-associated factor 3; Ub—ubiquitination. This figure was created with biorender.com.

Figure 4.
Contribution of CD79a and CD79b to the BCR-related signaling in lymphomas as a component of the cancer-mutation network. Abbreviations: BAFF—B-cell activating factor; BAFFR—B-cell activating factor receptor; BCL10—B-cell lymphoma 10; BCR—B-cell receptor; CARD11—caspase recruitment domain family member 11; DLBCL—diffuse large B-cell lymphoma; ITAM—immunoreceptor tyrosine-based activation motif; ITIM—immunoreceptor tyrosine-based inhibitory motif; Lyn—Lck/Yes novel tyrosine kinase; MALT1—mucosa-associated lymphoid tissue lymphoma translocation protein 1; MYD88—myeloid differentiation primary response 88; NF-κB—nuclear factor kappa-light-chain-enhancer of activated B cells; PI3K—phosphoinositide 3-kinase; P—phosphate group; TLR9—toll-like receptor 9. This figure was created with biorender.com.
Figure 4.
Contribution of CD79a and CD79b to the BCR-related signaling in lymphomas as a component of the cancer-mutation network. Abbreviations: BAFF—B-cell activating factor; BAFFR—B-cell activating factor receptor; BCL10—B-cell lymphoma 10; BCR—B-cell receptor; CARD11—caspase recruitment domain family member 11; DLBCL—diffuse large B-cell lymphoma; ITAM—immunoreceptor tyrosine-based activation motif; ITIM—immunoreceptor tyrosine-based inhibitory motif; Lyn—Lck/Yes novel tyrosine kinase; MALT1—mucosa-associated lymphoid tissue lymphoma translocation protein 1; MYD88—myeloid differentiation primary response 88; NF-κB—nuclear factor kappa-light-chain-enhancer of activated B cells; PI3K—phosphoinositide 3-kinase; P—phosphate group; TLR9—toll-like receptor 9. This figure was created with biorender.com.


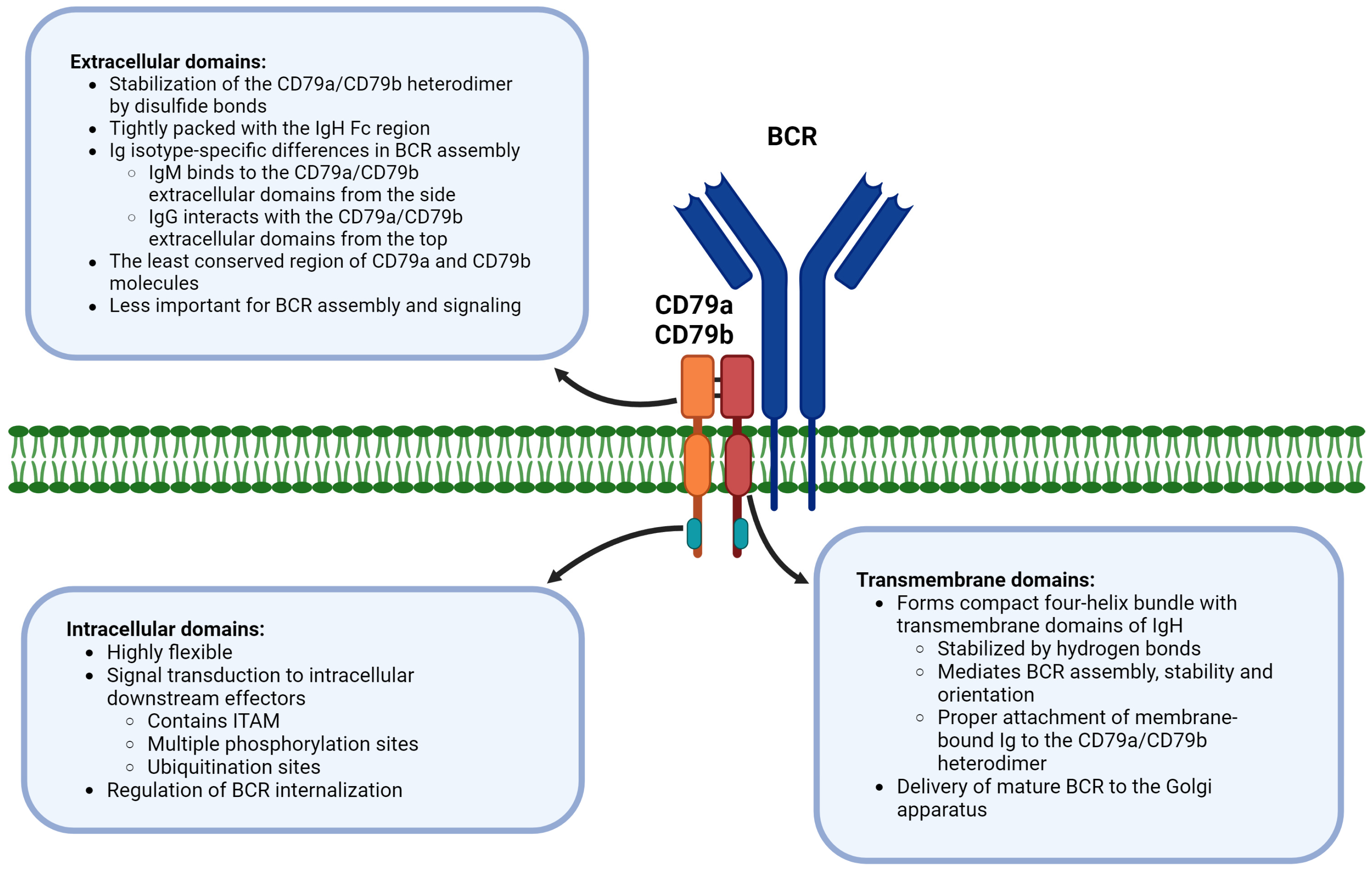{kind=link}
{kind=link}
{kind=link}
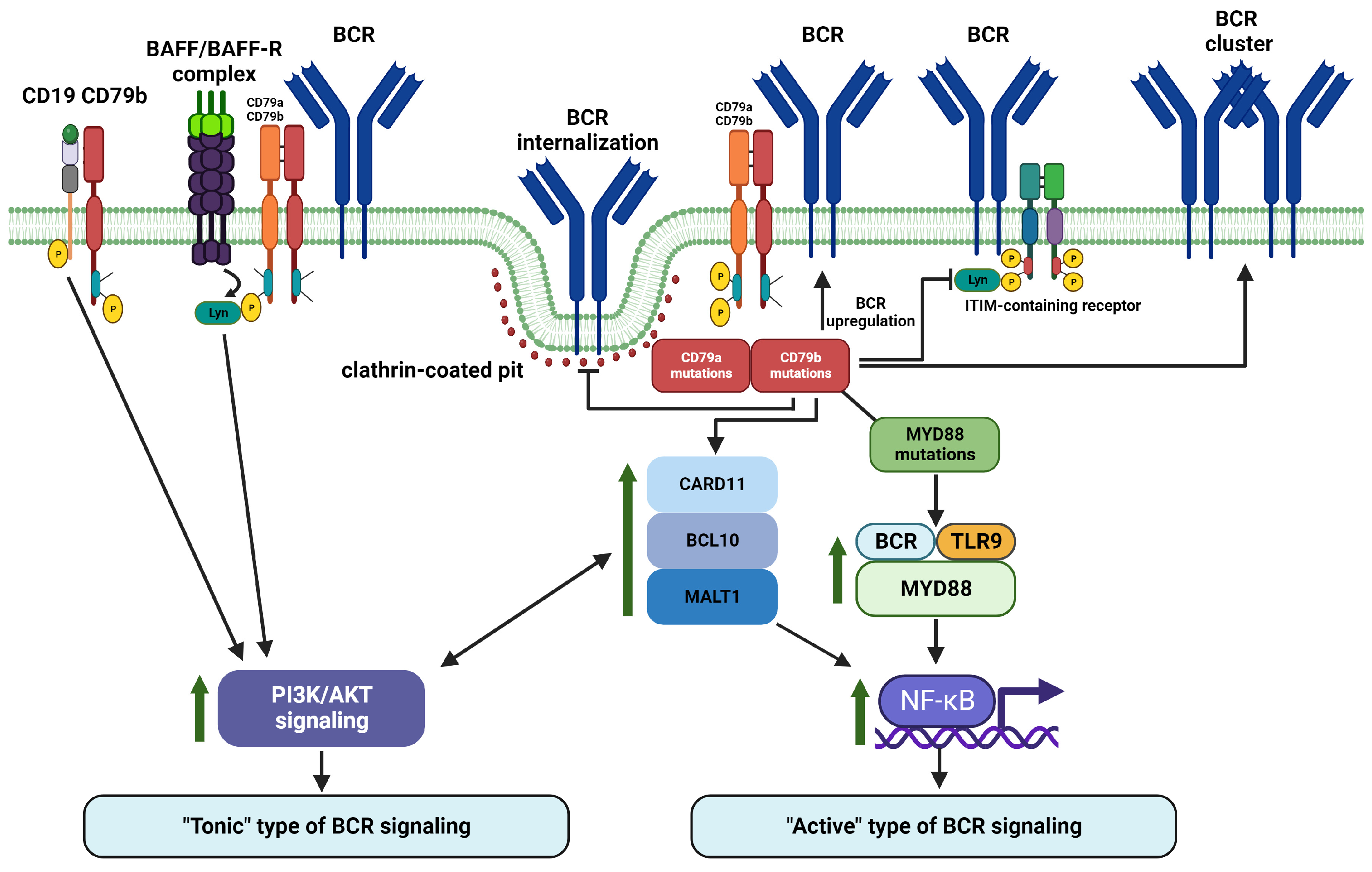{kind=link}
Table 1.
Frequency of CD79A mutations and related B-cell receptor signaling consequences.
| Frequency of Cases with Any CD79A ITAM Mutation | CD79A Mutation Subtypes | Frequency of Cases with CD79A Mutation Subtypes * | Signaling Consequences of CD79A Mutation Subtypes |
|---|---|---|---|
| 6% in DLBCL [124] 5% in unclassified DLBCL [125] 2.5% in CD5+ DLBCL [138] 0.7% in DLBCL [58] 6.5% in ABC DLBCL [139] 2.9% in ABC DLBCL [48] 0% in ABC DLBCL [125] 0% in GCB DLBCL [139] 6.3% in PCLBCL-LT [131] 5.5% in WM [140] 3% in FL [141] 3% in FL [124] 0% in FL [142] 0.8% in CLL [124] 0% in BL [124] 0% in SMZL [143] 0% in MZL [143] | CD79A ITAM Y188 point mutations | Not detected in multiple studies of DLBCL, FL, and other lymphoid malignancies | Inhibition of tonic BCR signaling, based on a cell line study [50] |
| CD79A ITAM Y199 point mutations | 1.8% in WM [140] | Not reported | |
| Complete ITAM deletions | 1.5% in ABC DLBCL [48] | Increased surface BCR expression [125] Chronic active BCR signal enhancement [139] | |
| Other CD79A mutations affecting ITAM (e.g., deletions, truncations, frameshift, or splice site mutations) | 6% in PCLBCL-LT (ITAM deletions affecting Y188) [131] 6.5% in ABC DLBCL [139] 1.3% in DLBCLs (deletions affecting Y188) [125] 2.9% in ABC DLBCL(splice site mutations) [48] 1.8% in WM (deletion) [140] | Increased surface BCR expression [125] Chronic active BCR signaling enhancement [139] |
* For studies with available information. Abbreviations: ABC DLBCL—activated B-cell-like diffuse large B-cell lymphoma; BL—Burkitt lymphoma; CLL—chronic lymphoblastic lymphoma; DLBCL—diffuse large B-cell lymphoma; FL—follicular lymphoma; GCB DLBCL—germinal center B-cell like diffuse large B-cell lymphoma; ITAM—immunoreceptor tyrosine-based activation motif; MZL—marginal zone lymphoma; NMZL—nodal marginal zone lymphoma; PCLBCL-LT—primary cutaneous large B-cell lymphoma, leg type; SMZL—splenic marginal zone lymphoma; WM—Waldenström macroglobulinemia.
Table 2.
Frequency of CD79B mutations and related B-cell receptor signaling consequences.
| Frequency of Cases with Any CD79B ITAM Mutation | CD79B Mutation Subtypes | Frequency of Cases with CD79B Mutation Subtypes * | Signaling Consequences of CD79B Mutation Subtypes |
|---|---|---|---|
| 65% in CLL [137,144] 56.3% in PCLBCL-LT [131] 33% in MZL [145] 0% in MZL [143] 30% in PCNSL [146] 31% in DLBCL [145] 38% in CD5+ DLBCLs [138] 26.8% in ABC DLBCL [57] 21.1% in ABC DLBCL [48] 10.8% in ABC DLBCL [147] 9% in ABC DLBCL [139] 23% in DLBCL [125] 14.5% in DLBCL [136] 14.3% in DLBCL [54] 14% in DLBCL [58] 12.9% in DLBCL [124] 8.5% in DLBCL [147] 8% in DLBCL [148] 5.6% in DLBCL [139] 5.1% in GCB DLBCL [147] 3.1% in GCB DLBCL [48] 1.9% in GCB DLBCL [57] 1.6% in GCB DLBCL [139] 10% in SMZL [143] 9% in WM [140] 7% in WM [149] 9% in FL [141] 5% in FL [124] 4.9% in FL [142] 0% in FL [148] 0% in FL [145] 0% in CLL [124] 0% in CLL [150] 0% in BL [48,124] 0%in MCL [148] 0% in MCL [145] 0% in marginal zone/MALT lymphoma [148] 0% gastric MALT lymphoma [48] 0% in SLL [148] 0% in CLL/SLL [145] 0% in NMZL [145] | CD79B ITAM Y196 point mutations | 75% in transformed WM [151] 53.1% in PCLBCL-LT [131] 30% in PCNSL [146] 35% in CD5+ DLBCL [138] 18% in ABC DLBCL [48] 1.6% in GCB DLBCL [48] 3.7% in DLBCL [139] 10% in SMZL [143] 9% in WM [140] 8% in DLBCL [148] 3% in FL [141] 7.5% in DLBCL [147] | Surface BCR expression upregulation via inhibition of BCR internalization [48,152] Reduced BCR signaling negative regulation via decreased Lyn binding [48] Enhancement of BCR clustering [3,50] Active BCR signaling enhancement [48] NF-κB activation enhancement [48] |
| CD79B ITAM Y207 point mutations | 4% in CLL [144,153] | Surface BCR expression upregulation [152] Impairment of antigen induced BCR signaling [144,153] | |
| Other CD79B mutations affecting ITAM (e.g., deletions, truncations, frameshift, splice site/intron retention mutations) | 0.7% in ABC DLBCLs (Y196 deletions) [48] 0.6% in DLBCL (deletions affecting Y207) [139] 0.5% in DLBCL (E197stop) [147] 17% in B-CLL (deletions) [144] 1.8% in DLBCL (splice site mutations and intron retention) [134] | Similar consequences as Y196 mutations | |
| CD79B ITAM mutations not affecting ITAM tyrosines | 2.5% in CD5+ DLBCL (deletion in ITAM region before the first tyrosine) [138] 1.6% in GCB DLBCLs (L199Q) [48] 6% in FL (point mutations in ITAM) [141] 3.1% in PCLBCL-LT (E197D point mutation) [131] 31% in DLBCL [145] 1.2% in DLBCLs [139] 12.7% in DLBCL (point mutations and one frameshift) [136] 25% in CLL [137] | Not reported | |
| Non-ITAM mutations | Over 30% in CLL [137] 17% in CLL [144] 1.8% in DLBCL [136] | Decreased BCR expression and signaling in CLL [153] |
* For studies with available information. Abbreviations: ABC DLBCL—activated B-cell like diffuse large B-cell lymphoma; BL—Burkitt lymphoma; CLL—chronic lymphoblastic lymphoma; DLBCL—diffuse large B-cell lymphoma; FL—follicular lymphoma; GCB DLBCL—germinal center B-cell like diffuse large B-cell lymphoma; ITAM—immunoreceptor tyrosine-based activation motif; MALT—mucosa-associated lymphatic tissue; MCL—mantle cell lymphoma; MZL—marginal zone lymphoma; NMZL—nodal marginal zone lymphoma; PCNSL—primary central nervous system lymphoma; PCLBCL-LT—primary cutaneous large B-cell lymphoma, leg type; SLL—small lymphocytic lymphoma; SMZL—splenic marginal zone lymphoma; WM—Waldenström macroglobulinemia.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tkachenko, A.; Kupcova, K.; Havranek, O. B-Cell Receptor Signaling and Beyond: The Role of Igα (CD79a)/Igβ (CD79b) in Normal and Malignant B Cells. Int. J. Mol. Sci. 2024, 25, 10. https://doi.org/10.3390/ijms25010010
AMA Style
Tkachenko A, Kupcova K, Havranek O. B-Cell Receptor Signaling and Beyond: The Role of Igα (CD79a)/Igβ (CD79b) in Normal and Malignant B Cells. International Journal of Molecular Sciences. 2024; 25(1):10. https://doi.org/10.3390/ijms25010010
Chicago/Turabian StyleTkachenko, Anton, Kristyna Kupcova, and Ondrej Havranek. 2024. "B-Cell Receptor Signaling and Beyond: The Role of Igα (CD79a)/Igβ (CD79b) in Normal and Malignant B Cells" International Journal of Molecular Sciences 25, no. 1: 10. https://doi.org/10.3390/ijms25010010
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.