
Characterisation of the RNA Virome of Nine Ochlerotatus Species in Finland

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
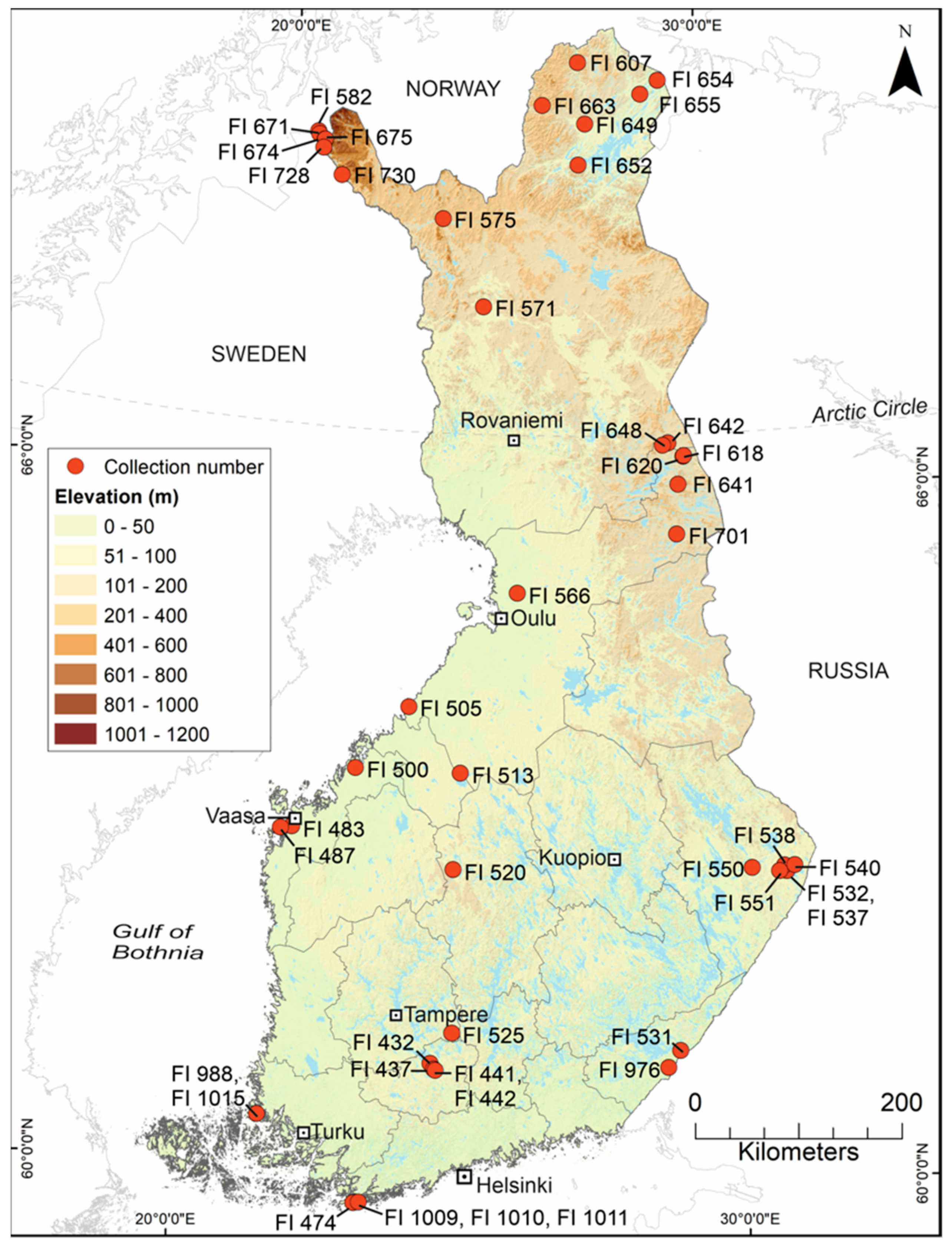
2.1. Mosquito Collection and Identification
2.2. Pooling and Homogenisation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection No. | Latitude (N) | Longitude (E) | Location-Pool No. | No. of Specimens | Collection Date | Mosquito Species |
|---|---|---|---|---|---|---|
| FI 432 | 61.0766 | 24.3912 | FIN/KH-2018/029 | 30 | 27 May 2015 | Oc. pullatus |
| FIN/KH-2018/047 | 20 | 27 May 2015 | Oc. punctor/punctodes | |||
| FI 437 | 61.0285 | 24.4596 | FIN/KH-2018/048 | 20 | 02 June 2015 | Oc. communis |
| FI 441 | 61.0201 | 24.4877 | FIN/KH-2018/038 | 13/20 | 02 June 2015 | Oc. intrudens |
| FIN/KH-2018/049 | 24 | 02 June 2015 | Oc. communis | |||
| FI 442 | 61.0223 | 24.4912 | FIN/KH-2018/038 | 7/20 | 02 June 2015 | Oc. intrudens |
| FI 474 | 59.8372 | 23.1595 | FIN/U-2018/050 | 20 | 14 June 2015 | Oc. communis |
| FI 483 | 63.0630 | 21.5680 | FIN/Po-2018/022 | 24 | 16 June 2015 | Oc. communis |
| FI 487 | 63.0410 | 21.3539 | FIN/Po-2018/009 | 27 | 16 June 2015 | Oc. excrucians |
| FI 500 | 63.6071 | 22.7055 | FIN/Po-2018/031 | 20 | 17 June 2015 | Oc. communis |
| FI 505 | 64.1637 | 23.6876 | FIN/PP-2018/010 | 60 | 17 June 2015 | Oc. communis |
| FI 513 | 63.6039 | 24.7534 | FIN/KP-2018/032 | 25 | 18 June 2015 | Oc. communis |
| FIN/KP-2018/033 | 16 | 18 June 2015 | Oc. diantaeus | |||
| FIN/KP-2018/034 | 20 | 18 June 2015 | Oc. intrudens | |||
| FI 520 | 62.7665 | 24.6814 | FIN/KS-2018/035 | 24 | 18 June 2015 | Oc. communis |
| FI 525 | 61.3473 | 24.7655 | FIN/Pi-2018/051 | 20 | 19 June 2015 | Oc. communis |
| FIN/Pi-2018/052 | 20 | 19 June 2015 | Oc. communis | |||
| FIN/Pi-2018/053 | 20 | 19 June 2015 | Oc. communis | |||
| FIN/Pi-2018/054 | 20 | 19 June 2015 | Oc. communis | |||
| FIN/Pi-2018/055 | 21 | 19 June 2015 | Oc. communis | |||
| FI 531 | 61.2013 | 28.9019 | FIN/EK-2018/056 | 22 | 25 June 2015 | Oc. communis |
| FI 532 | 62.7189 | 31.0050 | FIN/PK-2018/041 | 9/24 | 25 June 2015 | Oc. hexodontus |
| FIN/PK-2018/057 | 20 | 25 June 2015 | Oc. intrudens | |||
| FIN/PK-2018/058 | 20 | 25 June 2015 | Oc. diantaeus | |||
| FIN/PK-2018/059 | 20 | 25 June 2015 | Oc. communis | |||
| FIN/PK-2018/060 | 20 | 25 June 2015 | Oc. communis | |||
| FIN/PK-2018/061 | 20 | 25 June 2015 | Oc. intrudens | |||
| FI 537 | 62.7189 | 31.0050 | FIN/PK-2018/011 | 60 | 26 June 2015 | Oc. punctor/punctodes |
| FIN/PK-2018/041 | 15/24 | 26 June 2015 | Oc. hexodontus | |||
| FIN/PK-2018/042 | 20 | 26 June 2015 | Oc. cantans | |||
| FIN/PK-2018/062 | 20 | 26 June 2015 | Oc. communis | |||
| FIN/PK-2018/063 | 20 | 26 June 2015 | Oc. diantaeus | |||
| FIN/PK-2018/064 | 20 | 26 June 2015 | Oc. diantaeus | |||
| FIN/PK-2018/065 | 20 | 26 June 2015 | Oc. intrudens | |||
| FIN/PK-2018/066 | 20 | 26 June 2015 | Oc. intrudens | |||
| FIN/PK-2018/067 | 20 | 26 June 2015 | Oc. punctor/punctodes | |||
| FIN/PK-2018/068 | 20 | 26 June 2015 | Oc. intrudens | |||
| FIN/PK-2018/069 | 20 | 26 June 2015 | Oc. intrudens | |||
| FIN/PK-2018/070 | 20 | 26 June 2015 | Oc. communis | |||
| FIN/PK-2018/071 | 18 | 26 June 2015 | Oc. punctor/punctodes | |||
| FI 538 | 62.7700 | 30.9733 | FIN/PK-2018/072 | 20 | 26 June 2015 | Oc. intrudens |
| FIN/PK-2018/073 | 20 | 26 June 2015 | Oc. intrudens | |||
| FI 540 | 62.7666 | 31.1629 | FIN/PK-2018/021 | 24 | 26 June 2015 | Oc. communis |
| FI 550 | 62.7650 | 30.3541 | FIN/PK-2018/036 | 20 | 27 June 2015 | Oc. communis |
| FIN/PK-2018/074 | 20 | 27 June 2015 | Oc. communis | |||
| FIN/PK-2018/075 | 20 | 27 June 2015 | Oc. intrudens | |||
| FIN/PK-2018/076 | 20 | 27 June 2015 | Oc. communis | |||
| FIN/PK-2018/077 | 20 | 27 June 2015 | Oc. communis | |||
| FIN/PK-2018/078 | 20 | 27 June 2015 | Oc. communis | |||
| FIN/PK-2018/079 | 20 | 27 June 2015 | Oc. communis | |||
| FI 551 | 62.7241 | 30.8721 | FIN/PK-2018/080 | 21 | 27 June 2015 | Oc. intrudens |
| FI 566 | 65.1798 | 25.8002 | FIN/PP-2018/020 | 16 | 03 July 2015 | Oc. diantaeus |
| FI 571 | 67.6588 | 24.9049 | FIN/L-2018/008 | 48 | 03 July 2015 | Oc. intrudens |
| FI 575 | 68.4076 | 23.8850 | FIN/L-2018/005 | 32/48 | 04 July 2015 | Oc. communis |
| FIN/L-2018/027 | 8/24 | 04 July 2015 | Oc. communis | |||
| FI 582 | 69.0870 | 20.7600 | FIN/L-2018/005 | 8/48 | 02 July 2015 | Oc. communis |
| FI 607 | 69.7904 | 27.0549 | FIN/L-2018/001 | 48 | 07 July 2015 | Oc. hexodontus |
| FIN/L-2018/006 | 48 | 07 July 2015 | Oc. communis | |||
| FI 618 | 66.3588 | 29.3260 | FIN/PP-2018/015 | 40/57 | 09 July 2015 | Oc. punctor/punctodes |
| FIN/PP-2018/28 | 20 | 09 July 2015 | Oc. intrudens | |||
| FI 620 | 66.3639 | 29.3429 | FIN/PP-2018/015 | 17/57 | 09 July 2015 | Oc. punctor/punctodes |
| FIN/PP-2018/016 | 60 | 09 July 2015 | Oc. communis | |||
| FI 641 | 66.1148 | 29.1976 | FIN/PP-2018/082 | 20 | 18 July 2015 | Oc. communis |
| FIN/PP-2018/083 | 17 | 18 July 2015 | Oc. communis | |||
| FI 642 | 66.4756 | 29.0116 | FIN/L-2018/024 | 10/24 | 19 July 2015 | Oc. communis |
| FI 648 | 66.4597 | 28.8963 | FIN/L-2018/024 | 14/24 | 19 July 2015 | Oc. communis |
| FI 649 | 69.2558 | 27.2301 | FIN/L-2018/007 | 40/48 | 22 July 2015 | Oc. excrucians |
| FIN/L-2018/084 | 24 | 22 July 2015 | Oc. excrucians | |||
| FIN/L-2018/085 | 20 | 22 July 2015 | Oc. hexodontus | |||
| FIN/L-2018/086 | 20 | 22 July 2015 | Oc. hexodontus | |||
| FI 652 | 68.9008 | 27.0658 | FIN/L-2018/023 | 8/16 | 22 July 2015 | Oc. pullatus |
| FI 654 | 69.6249 | 29.0415 | FIN/L-2018/019 | 4/16 | 23 July 2015 | Oc. diantaeus |
| FIN/L-2018/007 | 1/48 | 23 July 2015 | Oc. excrucians | |||
| FI 655 | 69.5095 | 28.5965 | FIN/L-2018/019 | 12/16 | 23 July 2015 | Oc. diantaeus |
| FIN/L-2018/007 | 7/48 | 23 July 2015 | Oc. excrucians | |||
| FI 663 | 69.4178 | 26.1809 | FIN/L-2018/088 | 21 | 24 July 2015 | Oc. communis |
| FI 671 | 69.0617 | 20.7936 | FIN/L-2018/002 | 48 | 26 July 2015 | Oc. hexodontus |
| FIN/L-2018/003 | 48 | 26 July 2015 | Oc. punctor/punctodes | |||
| FIN/L-2018/026 | 24 | 26 July 2015 | Oc. punctor/punctodes | |||
| FI 674 | 69.0205 | 20.9304 | FIN/L-2018/089 | 20 | 28 July 2015 | Oc. hexodontus |
| FIN/L-2018/090 | 20 | 28 July 2015 | Oc. hexodontus | |||
| FI 675 | 69.0227 | 20.9380 | FIN/L-2018/030 | 22 | 28 July 2015 | Oc. hexodontus |
| FI 701 | 65.6855 | 29.1345 | FIN/PP-2018/004 | 48 | 23 August 2015 | Oc. punctor/punctodes |
| FI 728 | 68.9490 | 20.9210 | FIN/L-2018/005 | 8/48 | 02 July 2015 | Oc. communis |
| FIN/L-2018/023 | 8/16 | 02 July 2015 | Oc. pullatus | |||
| FI 730 | 68.7270 | 21.4220 | FIN/L-2018/027 | 16/24 | 03 July 2015 | Oc. communis? |
| FI 976 | 61.0569 | 28.6785 | FIN/EK-2018/040 | 20 | 04 July 2017 | Oc. communis |
| 61.0569 | 28.6785 | FIN/EK-2018/091 | 20 | 04 July 2017 | Oc. communis | |
| FI 988 | 60.5481 | 21.3696 | FIN/VS-2018/017 | 60 | 11 July 2017 | Oc. caspius |
| FI 1009 | 59.8439 | 23.2466 | FIN/U-2018/092 | 20 | 22 August 2017 | Oc. caspius |
| FIN/U-2018/093 | 17 | 22 August 2017 | Oc. punctor/punctodes | |||
| FI 1010 | 59.8439 | 23.2466 | FIN/U-2018/018 | 60 | 22–23 August 2017 | Oc. caspius |
| FIN/U-2018/039 | 25 | 22–23 August 2017 | Oc. punctor/punctodes | |||
| FIN/U-2018/094 | 20 | 22–23 August 2017 | Oc. caspius | |||
| FIN/U-2018/095 | 20 | 22–23 August 2017 | Oc. caspius | |||
| FI 1011 | 59.8439 | 23.2466 | FIN/U-2018/044 | 20 | 23–24 August 2017 | Oc. caspius |
| FIN/U-2018/045 | 21 | 23–24 August 2017 | Oc. punctor/punctodes | |||
| FIN/U-2018/096 | 20 | 23–24 August 2017 | Oc. caspius | |||
| FIN/U-2018/097 | 19 | 23–24 August 2017 | Oc. caspius | |||
| FI 1015 | 60.5481 | 21.3696 | FIN/VS-2018/098 | 20 | 24 August 2017 | Oc. caspius |
| FIN/VS-2018/099 | 20 | 24 August 2017 | Oc. caspius | |||
| FIN/VS-2018/100 | 26 | 24 August 2017 | Oc. caspius |
2.3. Illumina MiSeq Sequencing
2.4. NGS Data Analysis
3. Results
3.1. RNA Viromes Obtained Directly from Mosquito Homogenates
3.1.1. Positive-Sense ssRNA Virus Sequences
| Virus Family/ Taxon | Virus Name | Pool/Variant No. | Associated Mosquito Species | GenBank Accession |
|---|---|---|---|---|
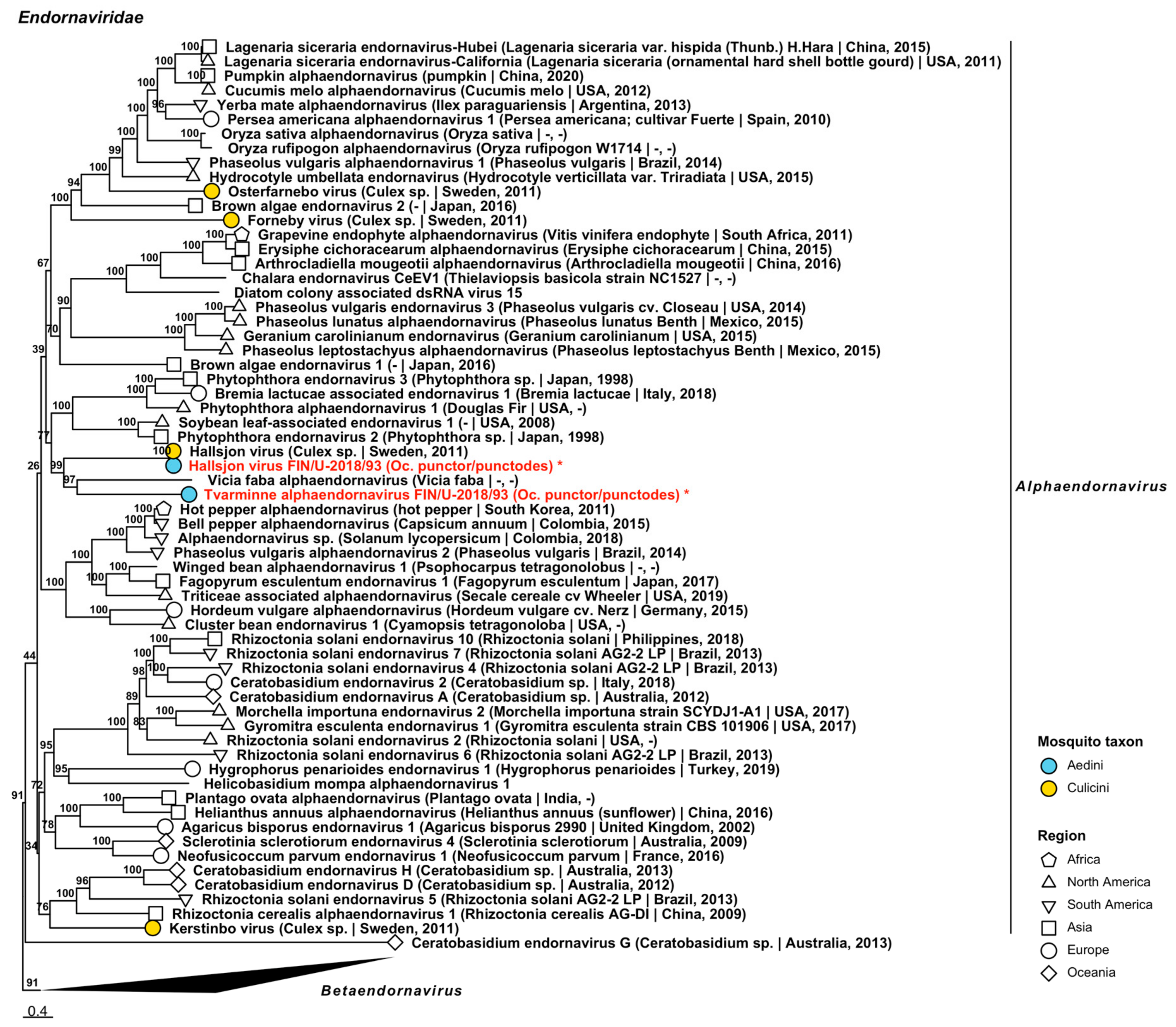
| Endornaviridae | Hallsjon virus | FIN/U-2018/93 | Oc. punctor/punctodes | ON955055 |
| Endornaviridae | Tvarminne alphaendornavirus | FIN/U-2018/93 | Oc. punctor/punctodes | ON955056 |
| Flaviviridae | Hameenlinna flavivirus | FIN/KH-2018/38 | Oc. intrudens | ON955057 |
| Flaviviridae | Kilpisjarvi flavivirus | FIN/L-2018/90 | Oc. hexodontus | ON949931 |
| Flaviviridae | Lestijarvi flavi-like virus | FIN/KP-2018/33 | Oc. diantaeus | ON955060 |
| Flaviviridae | Hanko virus | FIN/U-2018/94 FIN/U-2018/95 FIN/U-2018/96 FIN/U-2018/97 | Oc. caspius Oc. caspius Oc. caspius Oc. caspius | ON949927 ON949928 ON949929 ON949930 |
| Flaviviridae | Inari jingmenvirus | FIN/L-2018/30 FIN/L-2018/86 | Oc. hexodontus Oc. hexodontus | ON955058 ON955059 |
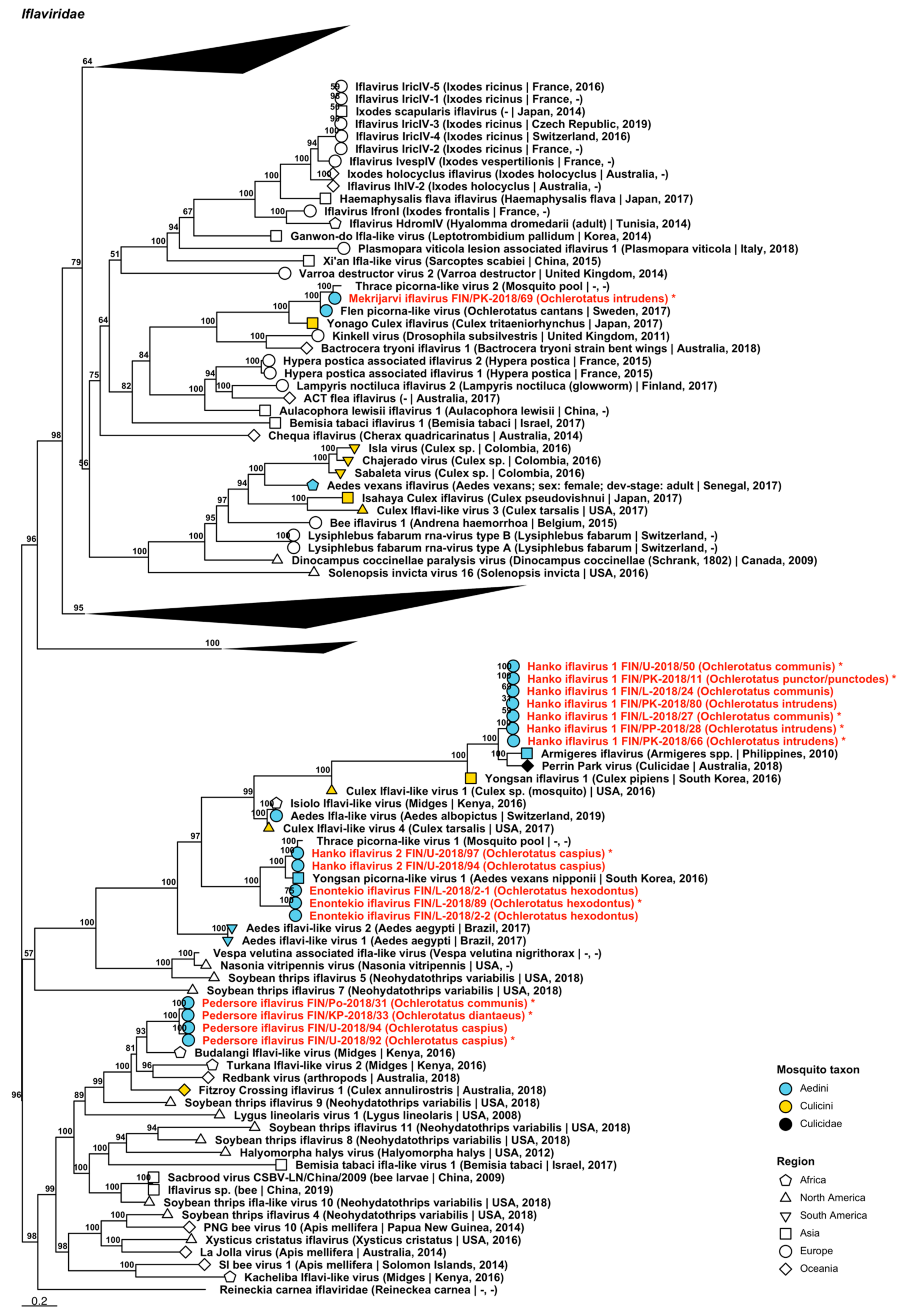
| Iflaviridae | Enontekio iflavirus | FIN/L-2018/02-1 FIN/L-2018/02-2 FIN/L-2018/89 | Oc. hexodontus Oc. hexodontus Oc. hexodontus | ON955061 ON955062 ON949932 |
| Iflaviridae | Hanko iflavirus 1 | FIN/PK-2018/11 FIN/L-2018/24 FIN/L-2018/27 FIN/PP-2018/28 FIN/U-2018/50 FIN/PK-2018/66 FIN/PK-2018/80 | Oc. punctor/punctodes Oc. communis Oc. communis Oc. intrudens Oc. communis Oc. intrudens Oc. intrudens | ON949934 ON955063 ON949933 ON949936 ON949937 ON949935 ON955064 |
| Iflaviridae | Hanko iflavirus 2 | FIN/U-2018/94 FIN/U-2018/97 | Oc. caspius Oc. caspius | ON955065 ON949938 |
| Iflaviridae | Mekrijarvi iflavirus | FIN/PK-2018/69 | Oc. intrudens | ON949939 |
| Iflaviridae | Pedersore iflavirus | FIN/Po-2018/31 FIN/KP-2018/33 FIN/U-2018/92 FIN/U-2018/94 | Oc. communis Oc. diantaeus Oc. caspius Oc. caspius | ON949941 ON949940 ON949942 ON955066 |
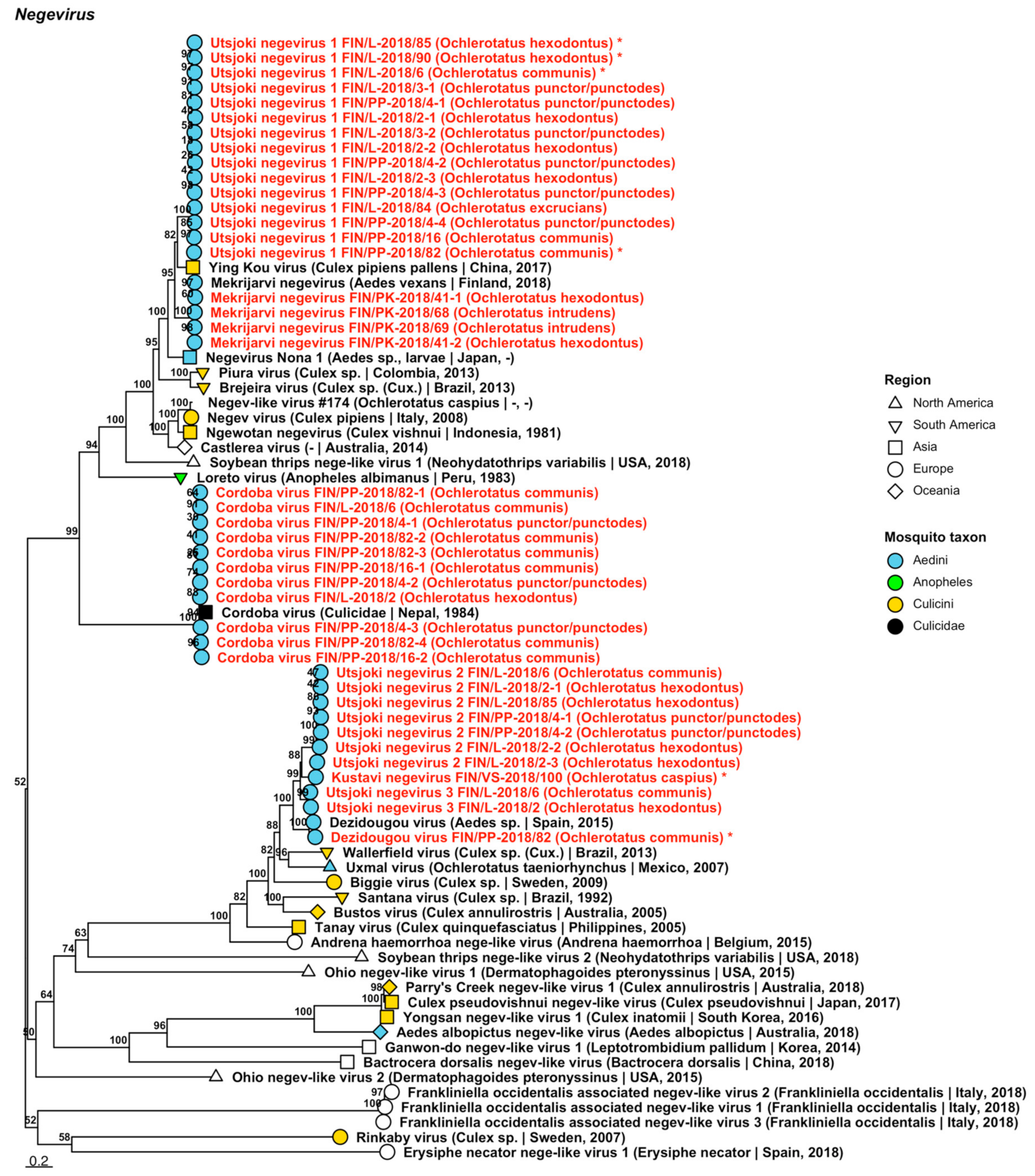
| Negevirus | Cordoba virus | FIN/L-2018/02 FIN/PP-2018/04-1 FIN/PP-2018/04-2 FIN/PP-2018/04-3 FIN/L-2018/06 FIN/PP-2018/16-1 FIN/PP-2018/16-2 FIN/PP-2018/82-1 FIN/PP-2018/82-2 FIN/PP-2018/82-3 FIN/PP-2018/82-4 | Oc. hexodontus Oc. punctor/punctodes Oc. punctor/punctodes Oc. punctor/punctodes Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis | ON955067 ON955069 ON955070 ON955071 ON955068 ON955072 ON955073 ON955074 ON955075 ON955076 ON955077 |
| Negevirus | Dezidougou virus | FIN/PP-2018/82 | Oc. communis | ON949943 |
| Negevirus | Kustavi negevirus | FIN/VS-2018/100 | Oc. caspius | ON949944 |
| Negevirus | Mekrijärvi negevirus | FIN/PK-2018/41-1 FIN/PK-2018/41-2 FIN/PK-2018/68 FIN/PK-2018/69 | Oc. hexodontus Oc. hexodontus Oc. intrudens Oc. intrudens | ON955078 ON955079 ON955080 ON955081 |
| Negevirus | Utsjoki negevirus 1 | FIN/L-2018/02-1 | Oc. hexodontus | ON955082 |
| FIN/L-2018/02-2 | Oc. hexodontus | ON955083 | ||
| FIN/L-2018/02-3 | Oc. hexodontus | ON955084 | ||
| FIN/L-2018/03-1 | Oc. punctor/punctodes | ON955085 | ||
| FIN/L-2018/03-2 | Oc. punctor/punctodes | ON955086 | ||
| FIN/PP-2018/04-1 | Oc. punctor/punctodes | ON955088 | ||
| Negevirus | Utsjoki negevirus 1 | FIN/PP-2018/04-2 FIN/PP-2018/04-3 FIN/PP-2018/04-4 FIN/U-2018/06 FIN/PP-2018/16 FIN/PP-2018/82 FIN/L-2018/84 FIN/L-2018/85 FIN/L-2018/90 | Oc. punctor/punctodes Oc. punctor/punctodes Oc. punctor/punctodes Oc. communis Oc. communis Oc. communis Oc. excrucians Oc. hexodontus Oc. hexodontus | ON955089 ON955090 ON955091 ON949945 ON955092 ON949948 ON955087 ON949946 ON949947 |
| Negevirus | Utsjoki negevirus 2 | FIN/L-2018/02-1 FIN/L-2018/02-2 FIN/L-2018/02-3 FIN/PP-2018/04-1 FIN/PP-2018/04-2 FIN/L-2018/06 FIN/L-2018/85 | Oc. hexodontus Oc. hexodontus Oc. hexodontus Oc. punctor/punctodes Oc. punctor/punctodes Oc. communis Oc. hexodontus | ON955093 ON955094 ON955095 ON955098 ON955099 ON955096 ON955097 |
| Negevirus | Utsjoki negevirus 3 | FIN/L-2018/02 FIN/L-2018/06 | Oc. hexodontus Oc. communis | ON955100 ON955101 |
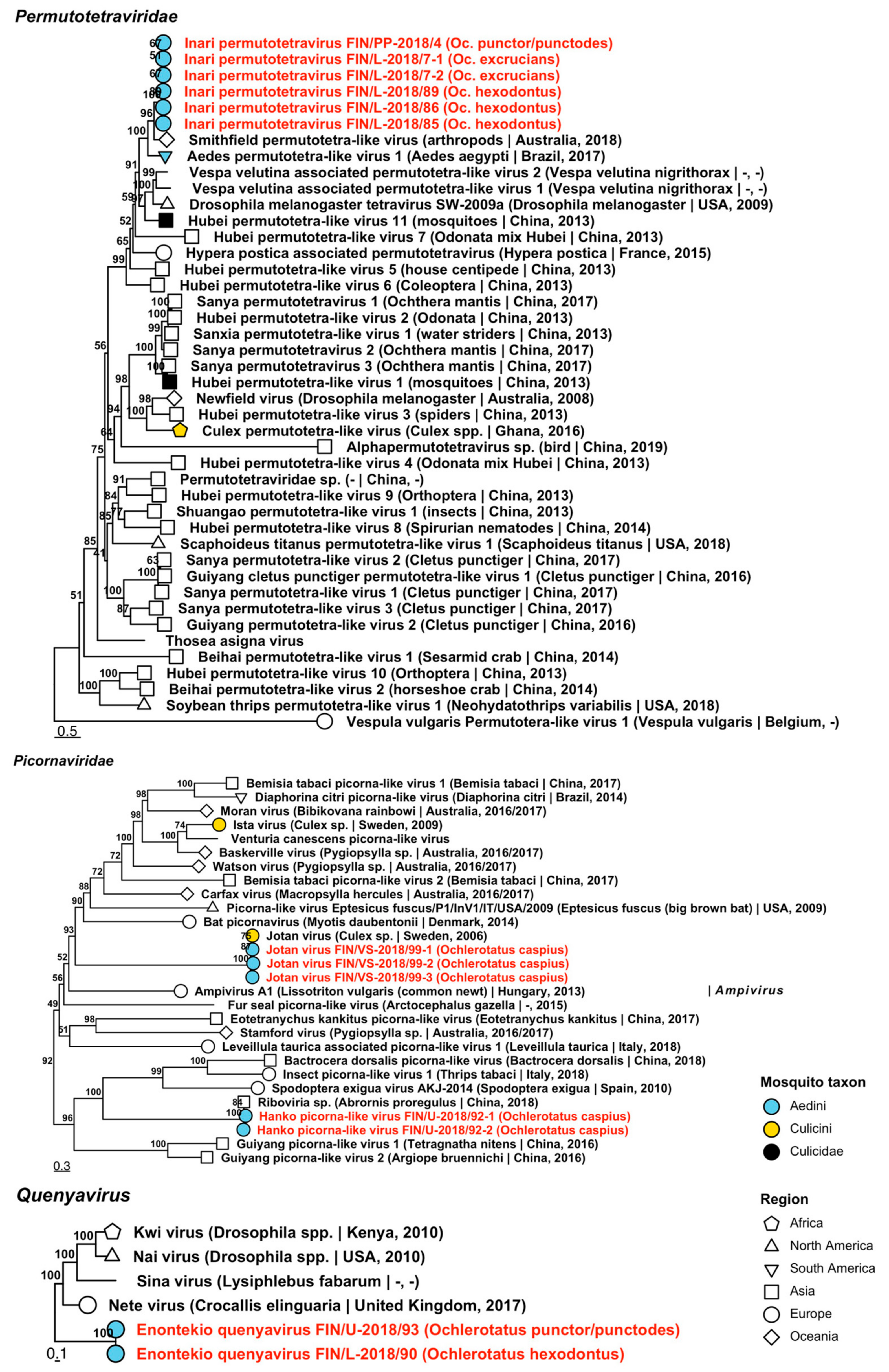
| Permutotetraviridae | Inari permutotetravirus | FIN/PP-2018/04 FIN/L-2018/07-1 FIN/L-2018/07-2 FIN/L-2018/85 FIN/L-2018/86 FIN/L-2018/89 | Oc. punctor/punctodes Oc. excrucians Oc. excrucians Oc. hexodontus Oc. hexodontus Oc. hexodontus | ON955107 ON955102 ON955103 ON955104 ON955105 ON955106 |
| Picornaviridae | Hanko picorna-like virus | FIN/U-2018/92-1 FIN/U-2018/92-2 | Oc. caspius Oc. caspius | ON955108 ON955109 |
| Picornaviridae | Jotan virus | FIN/VS-2018/99-1 FIN/VS-2018/99-2 FIN/VS-2018/99-3 | Oc. caspius Oc. caspius Oc. caspius | ON955110 ON955111 ON955112 |
| Quenyavirus | Enontekio quenyavirus | FIN/L-2018/90 FIN/U-2018/93 | Oc. hexodontus Oc. punctor/punctodes | ON955113 ON955114 |
| Solemoviridae | Enontekio sobemovirus | FIN/L-2018/02 FIN/L-2018/26 FIN/L-2018/89 | Oc. hexodontus Oc. punctor/punctodes Oc. hexodontus | ON955115 ON955116 ON955117 |
| Solemoviridae | Evros sobemo-like virus | FIN/VS-2018/17 FIN/U-2018/18 FIN/U-2018/92 FIN/U-2018/94 FIN/U-2018/95 FIN/U-2018/98 | Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius | ON955122 ON955118 ON955119 ON955120 ON955121 ON955123 |
| Solemoviridae | Hanko sobemovirus | FIN/U-2018/96 | Oc. caspius | ON955124 |
| Solemoviridae | Ilomantsi sobemovirus | FIN/L-2018/07 FIN/PK-2018/42 | Oc. excrucians Oc. cantans | ON955125 ON955126 |
| Solemoviridae | Joensuu sobemovirus | FIN/L-2018/19 FIN/PK-2018/75 FIN/PP-2018/82 | Oc. diantaeus Oc. intrudens Oc. communis | ON955127 ON955128 ON955129 |
| Togaviridae | Sindbis virus | FIN/PK-2018/62 | Oc. communis | ON955130 |
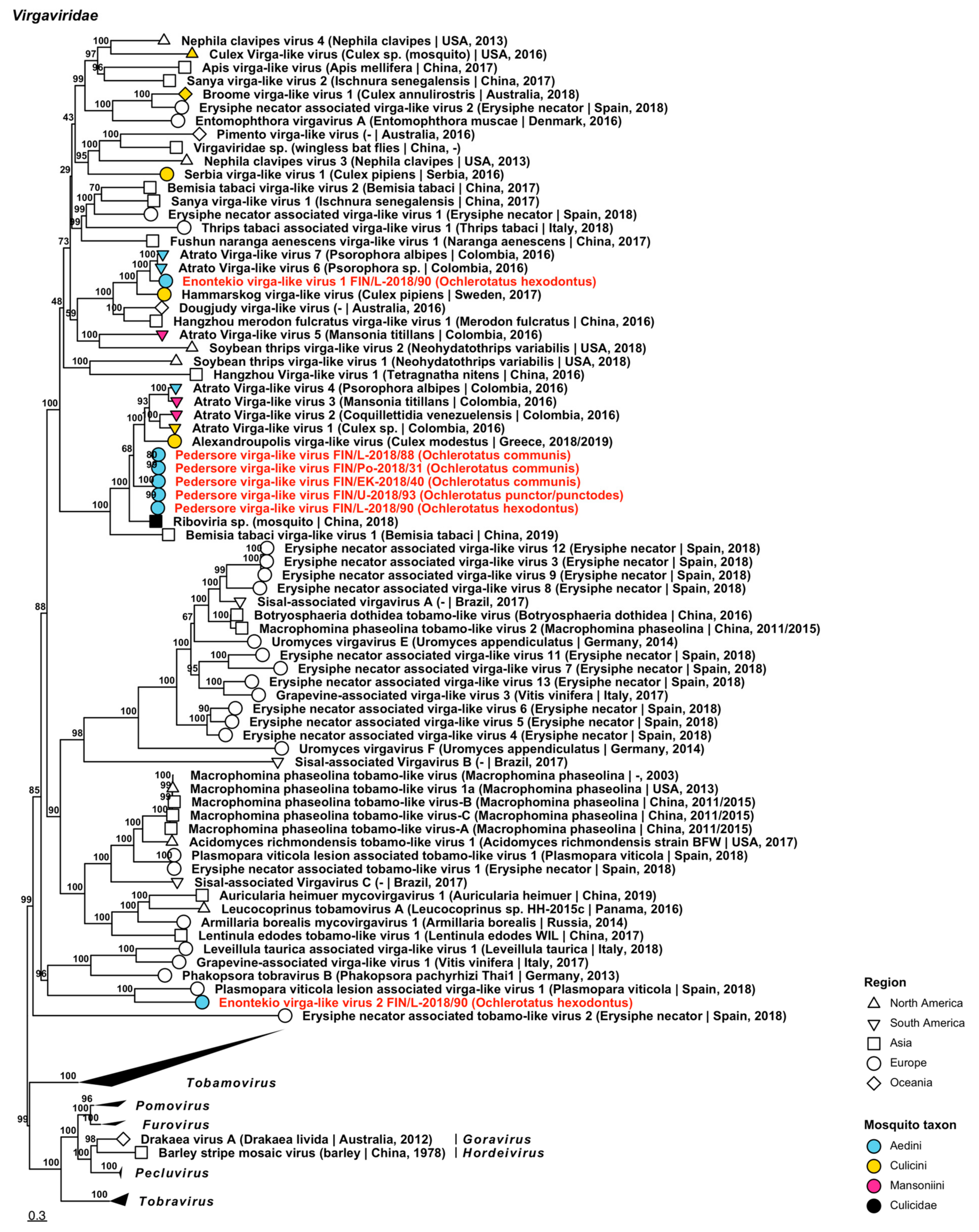
| Virgaviridae | Enontekio virga-like virus 1 | FIN/L-2018/90 | Oc. hexodontus | ON955131 |
| Virgaviridae | Enontekio virga-like virus 2 | FIN/L-2018/90 | Oc. hexodontus | ON955132 |
| Virgaviridae | Pedersore virga-like virus | FIN/Po-2018/31 FIN/EK-2018/40 FIN/L-2018/88 FIN/L-2018/90 FIN/U-2018/93 | Oc. communis Oc. communis Oc. communis Oc. hexodontus Oc. punctor/punctodes | ON955136 ON955133 ON955134 ON955135 ON955137 |





3.1.2. Negative-Sense ssRNA Virus Sequences

3.1.3. Negative-Sense ssRNA Virus Sequences
| Virus Family/ Taxon | Virus Name | Pool/Variant No. | Associated Mosquito Species | GenBank Accession |
|---|---|---|---|---|
| Aliusviridae | Lestijarvi obscuruvirus | FIN/KP-2018/32 | Oc. communis | ON955144 |
| Aspiviridae | Kilpisjarvi aspivirus | FIN/L-2018/90 | Oc. hexodontus | ON955145 |
| Chuviridae | Hattula chuvirus | FIN/L-2018/01-1 FIN/L-2018/01-2 FIN/L-2018/02 FIN/PP-2018/10-1 FIN/PP-2018/10-2 FIN/PP-2018/28-1 FIN/PP-2018/28-2 FIN/KH-2018/29 FIN/KP-2018/32 FIN/KS-2018/35 FIN/EK-2018/40 FIN/PK-2018/74 | Oc. hexodontus Oc. hexodontus Oc. hexodontus Oc. communis Oc. communis Oc. intrudens Oc. intrudens Oc. pullatus Oc. communis Oc. communis Oc. communis Oc. communis | ON955150 ON955151 ON955152 ON955154 ON955155 ON955156 ON955157 ON955147 ON955148 ON955149 ON955146 ON955153 |
| Chuviridae | Kustavi chuvirus 1 | FIN/VS-2018/17 | Oc. caspius | ON955158 |
| Chuviridae | Kustavi chuvirus 2 | FIN/VS-2018/17 | Oc. caspius | ON955159 |
| Phasmaviridae | Hameenlinna orthophasmavirus 1 | FIN/EK-2018/40 FIN/KH-2018/48 FIN/Pi-2018/51 FIN/Pi-2018/52 | Oc. communis Oc. communis Oc. communis Oc. communis | ON955160 ON955161 ON955162 ON955163 |
| Phasmaviridae | Hameenlinna orthophasmavirus 2 | FIN/EK-2018/40 FIN/KH-2018/48 | Oc. communis Oc. communis | ON955164 ON955165 |
| Phasmaviridae | Kuusamo orthophasmavirus 1 | FIN/PP-2018/83 | Oc. communis | ON955166 |
| Phasmaviridae | Kuusamo orthophasmavirus 2 | FIN/PP-2018/83 | Oc. communis | ON955167 |
| Phasmaviridae | Kuusamo orthophasmavirus 3 | FIN/PP-2018/83 | Oc. communis | ON955168 |
| Phasmaviridae | Kuusamo orthophasmavirus 4 | FIN/EK-2018/40 FIN/PP-2018/83 | Oc. communis Oc. communis | ON955169 ON955170 |
| Phasmaviridae | Lestijarvi orthophasmavirus 1 | FIN/KP-2018/34 | Oc. intrudens | ON955171 |
| Phasmaviridae | Lestijarvi orthophasmavirus 2 | FIN/KP-2018/34 | Oc. intrudens | ON955172 |



| Virus Family/ Taxon | Virus Name | Pool/Variant No. | Associated Mosquito Species | GenBank Accession |
|---|---|---|---|---|
| Phenuiviridae | Hameenlinna phasivirus | FIN/KP-2018/34-1 FIN/KP-2018/34-2 FIN/KP-2018/34-3 FIN/KH-2018/38-1 FIN/KH-2018/38-2 FIN/KH-2018/38-3 FIN/PK-2018/57-1 FIN/PK-2018/57-2 FIN/PK-2018/57-3 FIN/PK-2018/58-1 FIN/PK-2018/58-2 FIN/PK-2018/61 FIN/PK-2018/65-1 FIN/PK-2018/65-2 FIN/PK-2018/66-1 FIN/PK-2018/66-2 FIN/PK-2018/68 FIN/PK-2018/69-1 FIN/PK-2018/69-2 FIN/PK-2018/69-3 FIN/PK-2018/73-1 FIN/PK-2018/73-2 FIN/PK-2018/73-3 FIN/PK-2018/73-4 FIN/PK-2018/74-1 FIN/PK-2018/74-2 FIN/PK-2018/74-3 FIN/PK-2018/74-4 FIN/PK-2018/75-1 FIN/PK-2018/75-2 FIN/PK-2018/77-1 FIN/PK-2018/77-2 FIN/PK-2018/80-1 FIN/PK-2018/80-2 FIN/PK-2018/80-3 | Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. diantaeus Oc. diantaeus Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. communis Oc. communis Oc. communis Oc. communis Oc. intrudens Oc. intrudens Oc. communis Oc. communis Oc. intrudens Oc. intrudens Oc. intrudens | ON955181 ON955182 ON955183 ON955178 ON955179 ON955180 ON955184 ON955185 ON955186 ON955187 ON955188 ON955189 ON955190 ON955191 ON955192 ON955193 ON955138 ON955194 ON955195 ON955196 ON955197 ON955198 ON955199 ON955200 ON955201 ON955202 ON955203 ON955204 ON955205 ON955206 ON955207 ON955208 ON955209 ON955210 ON955211 |
| Phenuiviridae | Enontekio phenui-like virus 1 | FIN/L-2018/90 | Oc. hexodontus | ON955173 |
| Phenuiviridae | Enontekio phenui-like virus 2 | FIN/EK-2018/91 | Oc. communis | ON955174 |
| Phenuiviridae | Enontekio phenui-like virus 3 | FIN/L-2018/90 | Oc. hexodontus | ON955175 |
| Phenuiviridae | Enontekio phenui-like virus 4 | FIN/U-2018/93 | Oc. punctor/punctodes | ON955176 |
| Phenuiviridae | Enontekio phenui-like virus 5 | FIN/L-2018/90 | Oc. hexodontus | ON955177 |
| Phenuiviridae | Hanko phenui-like virus 1 | FIN/U-2018/96 | Oc. caspius | ON955212 |
| Phenuiviridae | Hanko phenui-like virus 2 | FIN/U-2018/93 | Oc. punctor/punctodes | ON955213 |
| Phenuiviridae | Hanko phenui-like virus 3 | FIN/U-2018/93 | Oc. punctor/punctodes | ON955214 |
| Phenuiviridae | Ilomantsi phenui-like virus | FIN/PK-2018/62 | Oc. communis | ON955215 |
| Phenuiviridae | Kalajoki phenui-like virus 1 | FIN/PP-2018/10 FIN/PK-2018/21 FIN/KP-2018/34-1 FIN/KP-2018/34-2 FIN/PK-2018/62 FIN/PK-2018/70 | Oc. communis Oc. communis Oc. intrudens Oc. intrudens Oc. communis Oc. communis | ON955221 ON955218 ON955216 ON955217 ON955219 ON955220 |
| Phenuiviridae | Kalajoki phenui-like virus 2 | FIN/PP-2018/10 FIN/KH-2018/48 FIN/Pi-2018/51 FIN/PK-2018/59 | Oc. communis Oc. communis Oc. communis Oc. communis | ON955225 ON955222 ON955223 ON955224 |
| Phenuiviridae | Palkane phenui-like virus 1 | FIN/Pi-2018/55 | Oc. communis | ON955226 |
| Phenuiviridae | Palkane phenui-like virus 2 | FIN/Pi-2018/52 FIN/Pi-2018/53 FIN/EK-2018/91 | Oc. communis Oc. communis Oc. communis | ON955228 ON955229 ON955227 |
| Qinviridae | Ilomantsi qinvirus | FIN/PK-2018/62 | Oc. communis | ON955230 |
| Qinviridae | Kalajoki qinvirus | FIN/PP-2018/10 FIN/Pi-2018/54 FIN/PK-2018/60 | Oc. communis Oc. communis Oc. communis | ON955233 ON955231 ON955232 |
| Qinviridae | Palkane qinvirus | FIN/Pi-2018/54 FIN/PK-2018/60-1 FIN/PK-2018/60-2 | Oc. communis Oc. communis Oc. communis | ON955234 ON955235 ON955236 |
| Rhabdoviridae | Enontekio merhavirus | FIN/L-2018/90 | Oc. hexodontus | ON955141 |
| Rhabdoviridae | Enontekio ohlsrhavirus | FIN/L-2018/30-1 FIN/L-2018/30-2 FIN/L-2018/30-3 FIN/L-2018/89 | Oc. hexodontus Oc. hexodontus Oc. hexodontus Oc. hexodontus | ON955237 ON955238 ON955239 ON955240 |
| Rhabdoviridae | Enontekio rhabdovirus | FIN/L-2018/03 | Oc. punctor/punctodes | ON955241 |
| Rhabdoviridae | Hattula rhabdovirus | FIN/KH-2018/29 FIN/KS-2018/35-1 FIN/KS-2018/35-2 FIN/PK-2018/59-1 FIN/PK-2018/59-2 FIN/PK-2018/62 FIN/PK-2018/76-1 FIN/PK-2018/76-2 FIN/L-2018/86-1 FIN/L-2018/86-2 | Oc. pullatus Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. hexodontus Oc. hexodontus | ON955242 ON955243 ON955244 ON955247 ON955248 ON955142 ON955249 ON955250 ON955245 ON955246 |
| Rhabdoviridae | Inari rhabdovirus | FIN/L-2018/84 | Oc. excrucians | ON955143 |
| Rhabdoviridae | Joutseno rhabdovirus 1 | FIN/EK-2018/91 | Oc. communis | ON955251 |
| Rhabdoviridae | Joutseno rhabdovirus 2 | FIN/EK-2018/91 | Oc. communis | ON955252 |
| Rhabdoviridae | Ohlsdorf virus | FIN/L-2018/07 FIN/L-2018/84 | Oc. excrucians Oc. excrucians | ON955253 ON955254 |
| Xinmoviridae | Enontekio anphevirus 1 | FIN/L-2018/90 | Oc. hexodontus | ON955255 |
| Xinmoviridae | Enontekio anphevirus 2 | FIN/L-2018/90 | Oc. hexodontus | ON955256 |
| Xinmoviridae | Hanko anphevirus | FIN/U-2018/96 | Oc. caspius | ON955257 |
| Xinmoviridae | Joensuu anphevirus | FIN/PK-2018/74 FIN/PP-2018/82 FIN/PP-2018/83-1 FIN/PP-2018/83-2 FIN/U-2018/93-1 FIN/U-2018/93-2 | Oc. communis Oc. communis Oc. communis Oc. communis Oc. punctor/punctodes Oc. punctor/punctodes | ON955258 ON955259 ON955260 ON955261 ON955262 ON955263 |
| Yueviridae | Enontekio yuevirus | FIN/L-2018/90 | Oc. caspius | ON955264 |



3.1.4. Double-Stranded RNA Virus Sequences

| Virus Family/ Taxon | Virus Name | Pool/Strain No. | Associated Mosquito Species | GenBank Accession |
|---|---|---|---|---|
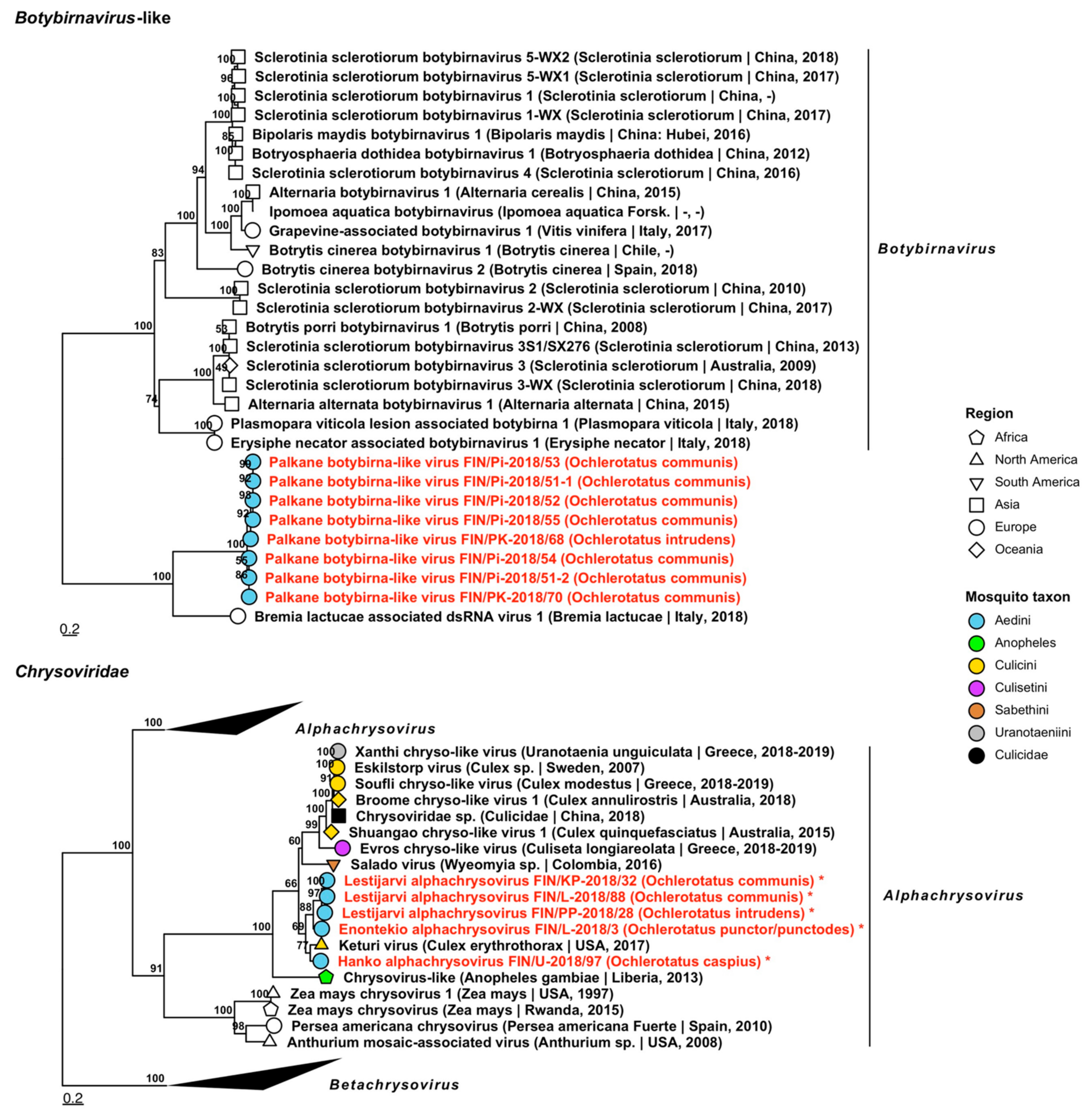
| Botybirnavirus | Palkane botybirna-like virus | FIN/Pi-2018/51-1 FIN/Pi-2018/51-2 FIN/Pi-2018/52 FIN/Pi-2018/53 FIN/Pi-2018/54 FIN/Pi-2018/55 FIN/PK-2018/68 FIN/PK-2018/70 | Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. intrudens Oc. communis | OP019912 OP019913 OP019914 OP019915 OP019916 OP019917 OP019918 OP019919 |
| Chrysoviridae | Enontekio alphachrysovirus | FIN/L-2018/03 | Oc. punctor/punctodes | OP019837–OP019840 |
| Chrysoviridae | Hanko alphachrysovirus | FIN/U-2018/97 | Oc. caspius | OP019841–OP019844 |
| Chrysoviridae | Lestijarvi alphachrysovirus | FIN/PP-2018/28 FIN/KP-2018/32 FIN/L-2018/88 | Oc. intrudens Oc. communis Oc. communis | OP019911, OP019846–OP019848 OP019910 OP019845 |

| Virus Family/ Taxon | Virus Name | Pool/Strain No. | Associated Mosquito Species | GenBank Accession |
|---|---|---|---|---|
| Partitiviridae | Enontekio alphapartitivirus 1 | FIN/L-2018/90 | Oc. hexodontus | OP019920 |
| Partitiviridae | Enontekio alphapartitivirus 2 | FIN/L-2018/90 | Oc. hexodontus | OP019921 |
| Partitiviridae | Hanko alphapartitivirus 1 | FIN/U-2018/94 FIN/U-2018/95 FIN/U-2018/96 | Oc. caspius Oc. caspius Oc. caspius | OP019929 OP019930 OP019931 |
| Partitiviridae | Hanko alphapartitivirus 2 | FIN/U-2018/96 | Oc. caspius | OP019932 |
| Partitiviridae | Hanko alphapartitivirus 3 | FIN/U-2018/93 | Oc. punctor/punctodes | OP019933 |
| Partitiviridae | Kalajoki alphapartitivirus | FIN/PP-2018/10 FIN/Pi-2018/52 FIN/PK-2018/68 | Oc. communis Oc. communis Oc. intrudens | OP019958 OP019956 OP019957 |
| Partitiviridae | Kuusamo alphapartitivirus | FIN/PP-2018/83 FIN/L-2018/90-1 FIN/L-2018/90-2 | Oc. communis Oc. hexodontus Oc. hexodontus | OP019963 OP019961 OP019962 |
| Partitiviridae | Palkane alphapartitivirus 1 | FIN/Pi-2018/51 FIN/Pi-2018/53 FIN/Pi-2018/55 | Oc. communis Oc. communis Oc. communis | OP019967 OP019968 OP019969 |
| Partitiviridae | Palkane alphapartitivirus 2 | FIN/Pi-2018/53 | Oc. communis | OP019970 |
| Partitiviridae | Enontekio betapartitivirus 1 | FIN/L-2018/90 | Oc. hexodontus | OP019922 |
| Partitiviridae | Enontekio betapartitivirus 2 | FIN/L-2018/90 | Oc. hexodontus | OP019923 |
| Partitiviridae | Kalajoki betapartitivirus | FIN/PP-2018/10 FIN/Pi-2018/51 | Oc. communis Oc. communis | OP019960 OP019959 |
| Partitiviridae | Ilomantsi deltapartitivirus | FIN/PP-2018/20 FIN/PK-2018/58 FIN/PK-2018/63 FIN/PK-2018/64 | Oc. diantaeus Oc. diantaeus Oc. diantaeus Oc. diantaeus | OP019944 OP019941 OP019942 OP019943 |
| Partitiviridae | Inari deltapartitivirus | FIN/L-2018/85 | Oc. hexodontus | OP019955 |
| Partitiviridae | Vaasa deltapartitivirus | FIN/L-2018/07 FIN/Po-2018/09 FIN/PK-2018/41 | Oc. excrucians Oc. excrucians Oc. hexodontus | OP019971 OP019972 OP019973 |
| Partitiviridae | Enontekio partiti-like virus | FIN/L-2018/23 | Oc. pullatus | OP019924 |
| Partitiviridae | Hattula partiti-like virus | FIN/L-2018/05 FIN/PP-2018/16 FIN/KH-2018/29 FIN/PK-2018/78 FIN/PP-2018/82 FIN/L-2018/86 FIN/EK-2018/91 | Oc. communis Oc. communis Oc. pullatus Oc. communis Oc. communis Oc. hexodontus Oc. communis | OP019936 OP019939 OP019935 OP019938 OP019940 OP019937 OP019934 |
| Partitiviridae | Hameenlinna partiti-like virus | FIN/PK-2018/42 FIN/KH-2018/48 FIN/U-2018/50 FIN/L-2018/88 | Oc. cantans Oc. communis Oc. communis Oc. communis | OP019927 OP019925 OP019928 OP019926 |
| Partitiviridae | Ilomantsi partiti-like virus 1 | FIN/L-2018/02 FIN/L-2018/08 FIN/PP-2018/15 FIN/PK-2018/72 FIN/L-2018/86 FIN/L-2018/89 FIN/L-2018/90 | Oc. hexodontus Oc. intrudens Oc. punctor/punctodes Oc. intrudens Oc. hexodontus Oc. hexodontus Oc. hexodontus | OP019945 OP019946 OP019951 OP019950 OP019947 OP019948 OP019949 |
| Partitiviridae | Ilomantsi partiti-like virus 2 | FIN/PK-2018/67 FIN/PK-2018/71 FIN/PK-2018/76 | Oc. punctor/punctodes Oc. punctor/punctodes Oc. communis | OP019952 OP019953 OP019954 |
| Partitiviridae | Kuusamo partiti-like virus | FIN/PP-2018/82 | Oc. communis | OP019964 |
| Partitiviridae | Lestijarvi partiti-like virus | FIN/KP-2018/34 FIN/PK-2018/41 | Oc. intrudens Oc. hexodontus | OP019965 OP019966 |
| Partitiviridae | Vaasa partiti-like virus | FIN/Po-2018/09 | Oc. excrucians | OP019974 |



| Virus Family/ Taxon | Virus Name | Pool/Strain No. | Associated Mosquito Species | GenBank Accession |
|---|---|---|---|---|
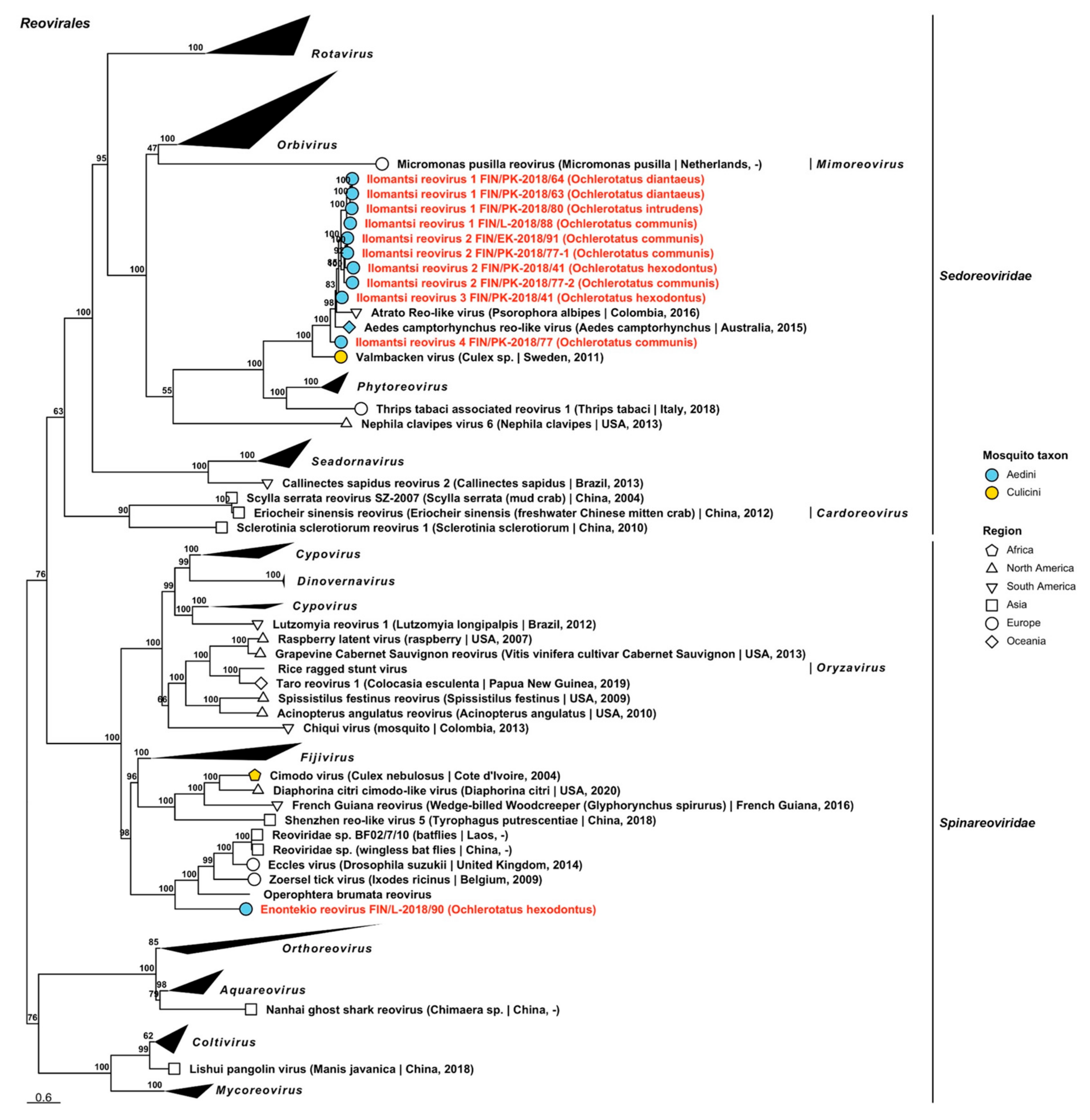
| Sedoreoviridae | Ilomantsi reovirus 1 | FIN/PK-2018/63 FIN/PK-2018/64 FIN/PK-2018/80 FIN/L-2018/88 | Oc. diantaeus Oc. diantaeus Oc. intrudens Oc. communis | OP019977 OP019978 OP019979 OP019976 |
| Sedoreoviridae | Ilomantsi reovirus 2 | FIN/PK-2018/41 FIN/PK-2018/77-1 FIN/PK-2018/77-2 FIN/EK-2018/91 | Oc. hexodontus Oc. communis Oc. communis Oc. communis | OP019981 OP019982 OP019983 OP019980 |
| Sedoreoviridae | Ilomantsi reovirus 3 | FIN/PK-2018/41 | Oc. hexodontus | OP019984 |
| Sedoreoviridae | Ilomantsi reovirus 4 | FIN/PK-2018/77 | Oc. communis | OP019985 |
| Spinareoviridae | Enontekio reovirus | FIN/L-2018/90 | Oc. hexodontus | OP019975 |
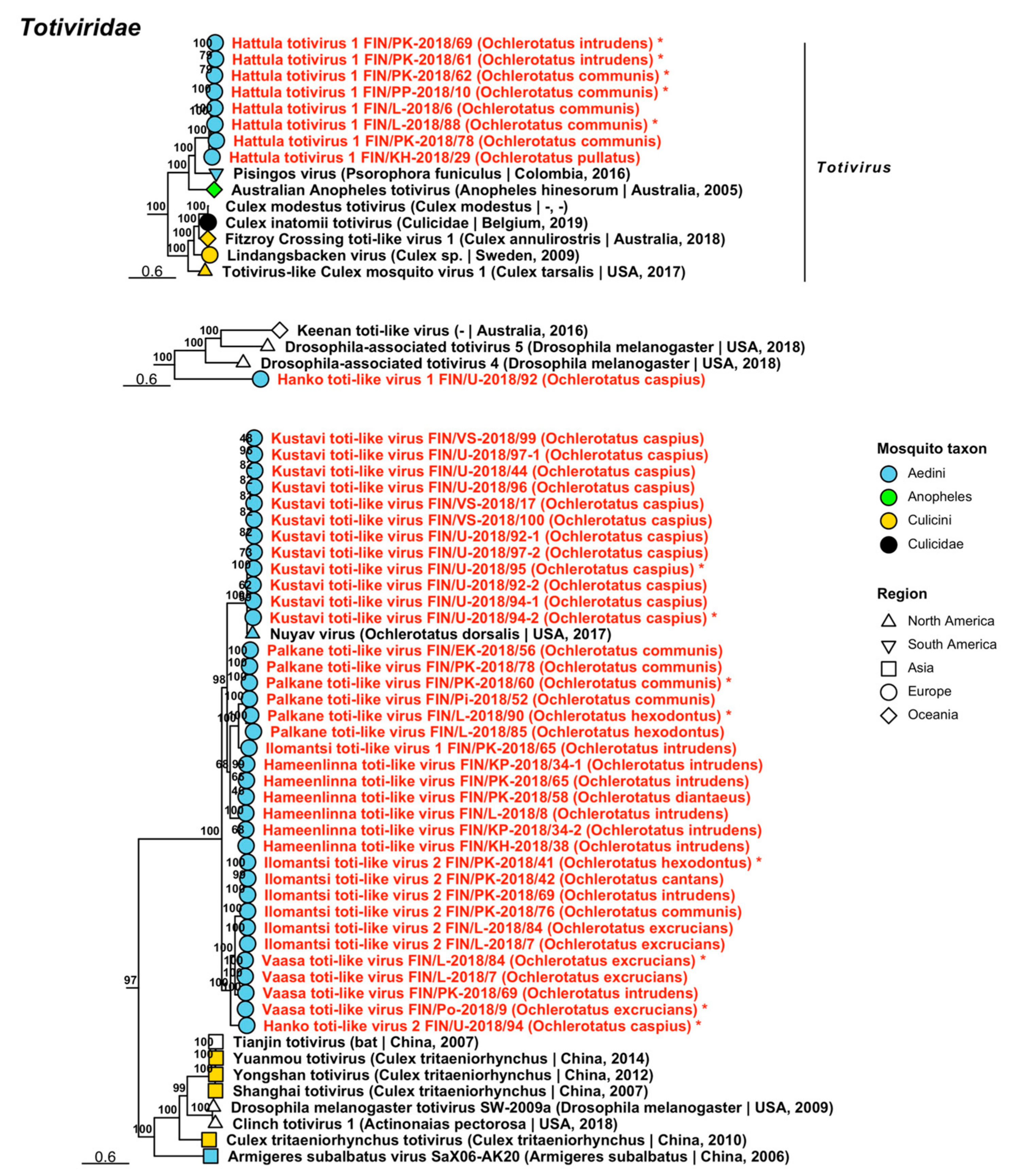
| Totiviridae | Hanko toti-like virus 1 | FIN/U-2018/92 | Oc. caspius | OP020048 |
| Totiviridae | Hanko toti-like virus 2 | FIN/U-2018/94 | Oc. caspius | OP019860 |
| Totiviridae | Hanko toti-like virus 3 | FIN/U-2018/93 | Oc. punctor/punctodes | OP020049 |
| Totiviridae | Enontekio toti-like virus 1 | FIN/L-2018/90 | Oc. hexodontus | OP019986 |
| Totiviridae | Enontekio toti-like virus 2 | FIN/L-2018/90 | Oc. hexodontus | OP019987 |
| Totiviridae | Enontekio toti-like virus 3 | FIN/L-2018/90 | Oc. hexodontus | OP019988 |
| Totiviridae | Enontekio toti-like virus 4 | FIN/L-2018/90 | Oc. hexodontus | OP019849 |
| Totiviridae | Enontekio totivirus 1 | FIN/L-2018/90 | Oc. hexodontus | OP019989 |
| Totiviridae | Enontekio totivirus 2 | FIN/L-2018/03 FIN/L-2018/90 | Oc. punctor/punctodes Oc. hexodontus | OP019990 OP019850 |
| Totiviridae | Enontekio totivirus 3 | FIN/L-2018/23 | Oc. pullatus | OP019991 |
| Totiviridae | Enontekio totivirus 4 | FIN/L-2018/23 | Oc. pullatus | OP019992 |
| Totiviridae | Enontekio totivirus 5 | FIN/L-2018/05 FIN/PP-2018/16 | Oc. communis Oc. communis | OP019993 OP019851 |
| Totiviridae | Enontekio totivirus 6 | FIN/L-2018/90 | Oc. hexodontus | OP019994 |
| Totiviridae | Enontekio totivirus 7 | FIN/L-2018/90 | Oc. hexodontus | OP019995 |
| Totiviridae | Hameenlinna toti-like virus | FIN/L-2018/08 FIN/KP-2018/34-1 FIN/KP-2018/34-2 FIN/KH-2018/38 FIN/PK-2018/58 FIN/PK-2018/65 | Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. diantaeus Oc. intrudens | OP019999 OP019997 OP019998 OP019996 OP020000 OP020001 |
| Totiviridae | Hameenlinna totivirus 1 | FIN/L-2018/05 FIN/PP-2018/10 FIN/PP-2018/16 FIN/PK-2018/21 FIN/Po-2018/31 FIN/PK-2018/36 FIN/EK-2018/40 FIN/KH-2018/48 FIN/KH-2018/49 FIN/U-2018/50 FIN/Pi-2018/51 FIN/Pi-2018/52-1 FIN/Pi-2018/52-2 FIN/Pi-2018/53 FIN/Pi-2018/54-1 FIN/Pi-2018/54-2 FIN/Pi-2018/55 FIN/EK-2018/56 FIN/PK-2018/59 FIN/PK-2018/60 FIN/PK-2018/68 FIN/PK-2018/69 FIN/PK-2018/70 FIN/PK-2018/74 FIN/PK-2018/76 FIN/PK-2018/78 FIN/PK-2018/79 FIN/PP-2018/82 FIN/PP-2018/83 FIN/L-2018/85-1 FIN/L-2018/85-2 FIN/L-2018/88 FIN/EK-2018/91 | Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. intrudens Oc. intrudens Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. hexodontus Oc. hexodontus Oc. communis Oc. communis | OP020006 OP020025 OP020026 OP020015 OP019856 OP020016 OP020002 OP020004 OP020005 OP020027 OP019854 OP020009 OP020010 OP020011 OP020012 OP020013 OP020014 OP020003 OP020017 OP020018 OP020019 OP020020 OP020021 OP020022 OP019855 OP020023 OP020024 OP019857 OP019858 OP020007 OP020008 OP019853 OP019852 |
| Totiviridae | Hameenlinna totivirus 2 | FIN/L-2018/08 FIN/KH-2018/38 FIN/PK-2018/75 FIN/PK-2018/80 | Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens | OP020029 OP020028 OP020030 OP020031 |
| Totiviridae | Hameenlinna totivirus 3 | FIN/L-2018/08-1 FIN/L-2018/08-2 FIN/KP-2018/34-1 FIN/KP-2018/34-2 FIN/KH-2018/38 FIN/PK-2018/57-1 FIN/PK-2018/57-2 FIN/PK-2018/61-1 FIN/PK-2018/61-2 FIN/PK-2018/65-1 FIN/PK-2018/65-2 FIN/PK-2018/68 FIN/PK-2018/73-1 FIN/PK-2018/73-2 FIN/PK-2018/75 FIN/PK-2018/80-1 FIN/PK-2018/80-2 | Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens Oc. intrudens | OP020034 OP020035 OP020032 OP020033 OP019859 OP020036 OP020037 OP020038 OP020039 OP020040 OP020041 OP020042 OP020043 OP020044 OP020045 OP020046 OP020047 |
| Totiviridae | Hanko toti-like virus 1 | FIN/U-2018/92 | Oc. caspius | OP020048 |
| Totiviridae | Hanko toti-like virus 2 | FIN/U-2018/94 | Oc. caspius | OP019860 |
| Totiviridae | Hanko toti-like virus 3 | FIN/U-2018/93 | Oc. punctor/punctodes | OP020049 |
| Totiviridae | Hanko totivirus 1 | FIN/U-2018/92 | Oc. caspius | OP020050 |
| Totiviridae | Hanko totivirus 2 | FIN/U-2018/92 | Oc. caspius | OP020052 |
| Totiviridae | Hanko totivirus 3 | FIN/U-2018/18 FIN/U-2018/44 FIN/U-2018/92 FIN/U-2018/94 FIN/U-2018/95 FIN/U-2018/96 FIN/U-2018/97 FIN/VS-2018/99 FIN/VS-2018/100 | Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius | OP020053 OP019861 OP019902 OP019909 OP019903 OP019904 OP019862 OP019905 OP020054 |
| Totiviridae | Hanko totivirus 4 | FIN/U-2018/18 FIN/U-2018/94 FIN/VS-2018/100 | Oc. caspius Oc. caspius Oc. caspius | OP020055 OP020056 OP020057 |
| Totiviridae | Hanko totivirus 5 | FIN/U-2018/18 FIN/U-2018/44 FIN/U-2018/92-1 FIN/U-2018/92-2 FIN/U-2018/92-3 FIN/U-2018/94 FIN/U-2018/95 FIN/U-2018/96 FIN/U-2018/97 FIN/VS-2018/99 FIN/VS-2018/100 | Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius | OP020058 OP020059 OP020060 OP020061 OP020062 OP020063 OP019906 OP020064 OP019863 OP020066 OP020065 |
| Totiviridae | Hanko totivirus 6 | FIN/U-2018/45 | Oc. punctor/punctodes | OP020067 |
| Totiviridae | Hanko totivirus 7 | FIN/U-2018/45 | Oc. punctor/punctodes | OP020068 |
| Totiviridae | Hanko totivirus 8 | FIN/U-2018/94 FIN/U-2018/95 | Oc. caspius Oc. caspius | OP019864 OP019865 |
| Totiviridae | Hanko totivirus 9 | FIN/U-2018/44 FIN/U-2018/94 FIN/U-2018/95 FIN/U-2018/96 | Oc. caspius Oc. caspius Oc. caspius Oc. caspius | OP020069 OP019866 OP019867 OP019900 |
| Totiviridae | Hanko totivirus 10 | FIN/U-2018/94 | Oc. caspius | OP020051 |
| Totiviridae | Hattula totivirus 1 | FIN/L-2018/06 FIN/PP-2018/10 FIN/KH-2018/29 FIN/PK-2018/61 FIN/PK-2018/62 FIN/PK-2018/69 FIN/PK-2018/78 FIN/L-2018/88 | Oc. communis Oc. communis Oc. pullatus Oc. intrudens Oc. communis Oc. intrudens Oc. communis Oc. communis | OP020071 OP019871 OP020070 OP019868 OP019869 OP019870 OP020072 OP019901 |
| Totiviridae | Hattula totivirus 2 | FIN/PP-2018/10 FIN/KH-2018/29-1 FIN/KH-2018/29-2 FIN/Po-2018/31 FIN/EK-2018/40-1 FIN/EK-2018/40-2 FIN/Pi-2018/52 FIN/Pi-2018/53 FIN/EK-2018/56 FIN/PK-2018/57 FIN/PK-2018/60 FIN/PK-2018/74 FIN/EK-2018/91 | Oc. communis Oc. pullatus Oc. pullatus Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis Oc. intrudens Oc. communis Oc. communis Oc. communis | OP019876 OP020075 OP020076 OP020080 OP020073 OP020074 OP020077 OP019874 OP019872 OP020078 OP019875 OP020079 OP019873 |
| Totiviridae | Hattula totivirus 3 | FIN/L-2018/03 FIN/L-2018/23 FIN/L-2018/26 FIN/U-2018/39 FIN/U-2018/45 FIN/KH-2018/47 FIN/PK-2018/60 FIN/PK-2018/62 FIN/PK-2018/66 FIN/L-2018/85 | Oc. punctor/punctodes Oc. pullatus Oc. punctor/punctodes Oc. punctor/punctodes Oc. punctor/punctodes Oc. punctor/punctodes Oc. communis Oc. communis Oc. intrudens Oc. hexodontus | OP020083 OP020081 OP020082 OP019881 OP019882 OP019877 OP019879 OP019880 OP020084 OP019878 |
| Totiviridae | Ilomantsi toti-like virus 1 | FIN/PK-2018/65 | Oc. intrudens | OP020085 |
| Totiviridae | Ilomantsi toti-like virus 2 | FIN/L-2018/07 FIN/PK-2018/41 FIN/PK-2018/42 FIN/PK-2018/69 FIN/PK-2018/76 FIN/L-2018/84 | Oc. excrucians Oc. hexodontus Oc. cantans Oc. intrudens Oc. communis Oc. excrucians | OP020086 OP019883 OP020088 OP020089 OP020090 OP020087 |
| Totiviridae | Ilomantsi toti-like virus 3 | FIN/PK-2018/58 | Oc. diantaeus | OP020091 |
| Totiviridae | Ilomantsi totivirus 1 | FIN/PK-2018/58 FIN/PP-2018/82 | Oc. diantaeus Oc. communis | OP020092 OP019884 |
| Totiviridae | Ilomantsi totivirus 2 | FIN/PK-2018/42-1 FIN/PK-2018/42-2 FIN/PK-2018/76 | Oc. cantans Oc. cantans Oc. communis | OP020093 OP020094 OP020095 |
| Totiviridae | Ilomantsi totivirus 3 | FIN/PK-2018/42 | Oc. cantans | OP020096 |
| Totiviridae | Inari toti-like virus | FIN/L-2018/84 FIN/U-2018/93 | Oc. excrucians Oc. punctor/punctodes | OP020097 OP020098 |
| Totiviridae | Inari totivirus 1 | FIN/L-2018/07-1 FIN/L-2018/07-2 FIN/L-2018/84-1 FIN/L-2018/84-2 | Oc. excrucians Oc. excrucians Oc. excrucians Oc. excrucians | OP020099 OP020100 OP019885 OP020101 |
| Totiviridae | Inari totivirus 2 | FIN/L-2018/19 FIN/L-2018/85 FIN/L-2018/88 | Oc. diantaeus Oc. hexodontus Oc. communis | OP019886 OP019887 OP019888 |
| Totiviridae | Joutseno totivirus | FIN/EK-2018/40 | Oc. communis | OP019889 |
| Totiviridae | Karstula totivirus | FIN/KS-2018/35 | Oc. communis | OP020102 |
| Totiviridae | Kustavi toti-like virus | FIN/VS-2018/17 FIN/U-2018/44 FIN/U-2018/92-1 FIN/U-2018/92-2 FIN/U-2018/94-1 FIN/U-2018/94-2 FIN/U-2018/95 FIN/U-2018/96 FIN/U-2018/97-1 FIN/U-2018/97-2 FIN/VS-2018/99 FIN/VS-2018/100 | Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius Oc. caspius | OP020111 OP020103 OP020104 OP020105 OP020106 OP019890 OP019891 OP020107 OP020108 OP020109 OP020112 OP020110 |
| Totiviridae | Kuusamo toti-like virus | FIN/PP-2018/83 | Oc. communis | OP020113 |
| Totiviridae | Kuusamo totivirus 1 | FIN/PP-2018/15 | Oc. punctor/punctodes | OP020114 |
| Totiviridae | Kuusamo totivirus 2 | FIN/PP-2018/15 | Oc. punctor/punctodes | OP020115 |
| Totiviridae | Lestijarvi totivirus | FIN/L-2018/19 FIN/KP-2018/33 FIN/PK-2018/58 FIN/PK-2018/63 FIN/PK-2018/64 FIN/PP-2018/82 | Oc. diantaeus Oc. diantaeus Oc. diantaeus Oc. diantaeus Oc. diantaeus Oc. communis | OP019892 OP020116 OP019893 OP020117 OP020118 OP019894 |
| Totiviridae | Palkane toti-like virus | FIN/Pi-2018/52 FIN/EK-2018/56 FIN/PK-2018/60 FIN/PK-2018/78 FIN/L-2018/85 FIN/L-2018/90 | Oc. communis Oc. communis Oc. communis Oc. communis Oc. hexodontus Oc. hexodontus | OP020121 OP020119 OP019896 OP020122 OP020120 OP019895 |
| Totiviridae | Palkane totivirus | FIN/EK-2018/40 FIN/Pi-2018/54-1 FIN/Pi-2018/54-2 FIN/Pi-2018/55 FIN/EK-2018/91 | Oc. communis Oc. communis Oc. communis Oc. communis Oc. communis | OP019907 OP020123 OP020124 OP020125 OP019908 |
| Totiviridae | Utsjoki toti-like virus | FIN/L-2018/88 | Oc. communis | OP019897 |
| Totiviridae | Vaasa toti-like virus | FIN/L-2018/07 FIN/Po-2018/09 FIN/PK-2018/69 FIN/L-2018/84 | Oc. excrucians Oc. excrucians Oc. intrudens Oc. excrucians | OP020126 OP019899 OP020127 OP019898 |
| Totiviridae | Vaasa totivirus | FIN/Po-2018/09 FIN/PK-2018/41 | Oc. excrucians Oc. hexodontus | OP020129 OP020128 |





3.2. Viruses by Mosquito Species
| Virus Family | No. Virus Variants | No. of Viruses | No. Novel Viruses | Host Associations | Oc. cantans (1) | Oc. caspius (11) | Oc. communis (35) | Oc. diantaeus (6) | Oc. excrucians (3) | Oc. hexodontus (8) | Oc. intrudens (14) | Oc. pullatus (2) | Oc. punctor/punctodes (11) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| + ssRNA | Endornaviridae | 2 | 2 | 1 | Plants, fungi and oomycetes; host-specific [39]. | - | - | - | - | - | - | - | - | 2/1 | No. of virus species per mosquito species/no. of novel virus species |
| Flaviviridae | 9 | 5 | 4 | Arthropod-borne; mammalian hosts [40] mosquitoes [7,8,9,41,42,43]. | - | 1 | - | 1/1 | - | 2/2 | 1/1 | - | - | ||
| Iflavirus | 17 | 5 | 5 | Arthropoda [44], mosquitoes, inc. Culex sp. [45]. | - | 2/2 | 2/2 | 1/1 | - | 1/1 | 2/2 | - | 1/1 | ||
| Negevirus | 41 | 7 | 4 | Phlebotomine sandflies and mosquitoes [10,46]. | - | 1/1 | 5/3 | - | 1/1 | 5/3 | 1 | - | 3/2 | ||
| Permutotetraviridae | 6 | 1 | 1 | Insecta: Setothosea asigna [47], Euprosterna elaeasa [48]. Fungus: Botrytis cinerea [49]. Mosquitoes [42]. | - | - | - | - | 1/1 | 1/1 | - | - | 1/1 | ||
| Picornaviridae | 5 | 2 | 0 | Vertebrates (six of the seven classes) [50], Culex mosquitoes [51] and fleas [52]. | - | 2 | - | - | - | - | - | - | - | ||
| Quenyavirus | 2 | 1 | 1 | Insecta: Crocallis elinguaria, Drosophila sp. and Lysiphlebus fabarum [53]. | - | - | - | - | - | 1/1 | - | - | 1/1 | ||
| Solemoviridae | 15 | 5 | 4 | Plants (monocotyledons and dicotyledons) [54]. | 1/1 | 2/1 | 1/1 | 1/1 | 1/1 | 1/1 | 1/1 | - | 1/1 | ||
| Togaviridae | 1 | 1 | 0 | Humans and nonhuman primates, mosquitoes, amphibians, arthropods, birds, equids, pigs, reptiles, rodents, salmonids and sea mammals; most are mosquito-borne [55]. Mosquitoes [43]. | - | - | 1 | - | - | - | - | - | - | ||
| Virgaviridae | 7 | 3 | 3 | Plants, plasmodiophorids, nematodes and pollen [56]. | - | - | 1/1 | - | - | 3/3 | - | - | 1/1 | ||
| − ssRNA | Aliusviridae | 1 | 1 | 1 | Mosquitoes, Coleoptera, Hymenoptera, leafhopper [57] fleas [52] | - | - | 1/1 | - | - | - | - | - | - | |
| Aspiviridae | 1 | 1 | 1 | Plants and plant-infecting fungi [58]. | - | - | - | - | - | 1/1 | - | - | - | ||
| Chuviridae | 14 | 3 | 3 | Mosquitoes [59,60], earwigs, Odonata, ticks, cockroaches, snakes, fish [61]. | - | 2/2 | 1/1 | - | - | 1/1 | 1/1 | 1/1 | - | ||
| Phasmaviridae | 13 | 8 | 8 | Mosquitoes [43], Hymenoptera, Hemiptera, Coleoptera [59]. | - | - | 6/6 | - | - | - | 2/2 | - | - | ||
| Phenuiviridae | 58 | 14 | 14 | Mosquitoes [43,62], fleas [52], ticks, Coleoptera, phlebotomine sandflies, plants, humans [61]. | - | 1/1 | 7/7 | 1/1 | - | 3/3 | 2/2 | - | 3/3 | ||
| Qinviridae | 7 | 3 | 3 | Insects [63,64], marine diatoms [57]. | - | - | 3/3 | - | - | - | - | - | - | ||
| Rhabdoviridae | 21 | 8 | 7 | Vertebrates, invertebrates and plants [61]. Insect vectors infect vertebrates [65], mosquitoes [32]. | - | - | 3/3 | - | 2/1 | 3/3 | - | 1/1 | 1/1 | ||
| Xinmoviridae | 9 | 4 | 4 | Mosquitoes [66,67], Odonata & Hymenoptera [68]. | - | 1/1 | 1/1 | - | - | 2/2 | - | - | 1/1 | ||
| Yueviridae | 1 | 1 | 1 | Invertebrates (freshwater isopoda/sesarmid crab) [35]. | - | - | - | - | - | 1/1 | - | - | - | ||
dsRNA | Botybirnavirus | 8 | 1 | 1 | Phytopathogenic fungi [69]. | - | - | 1/1 | - | - | - | 1/1 | - | - | |
| Chrysoviridae | 5 | 3 | 3 | Fungi and plants; possibly insects [70]. | - | 1/1 | 1/1 | - | - | - | 1/1 | - | 1/1 | ||
| Partitiviridae | 55 | 23 | 23 | Plants, fungi, protozoa [71], mosquitoes [60]. | 1/1 | 2/2 | 9/9 | 1/1 | 2/2 | 10/10 | 3/3 | 2/2 | 3/3 | ||
| Sedoreoviridae | 10 | 4 | 4 | Pathogenic viruses; arthropods [72,73,74], mammals, inc. humans [75,76], plants [77]. | - | - | 3/3 | 1/1 | - | 2/2 | 1/1 | - | - | ||
| Spinareoviridae | 1 | 1 | 0 | Pathogenic viruses; mosquitoes [78], plants [79], fish [79], reptiles, birds and mammals [80]. | - | - | - | - | - | 1/1 | - | - | - | ||
| Totiviridae | 205 | 52 | 50 | Fungi [81], protozoa [82], mosquitoes [60], fleas [52]. | 3/3 | 11/9 | 16/16 | 5/5 | 5/5 | 14/14 | 10/10 | 5/5 | 8/8 | ||
| Totals | 514 | 159 | 147 | 5 | 26 | 62 | 11 | 12 | 52 | 26 | 9 | 27 | |||
4. Discussion
4.1. Classification and Interpretation of the Viruses Detected in this Study
4.2. Reflections on the Methods and Their Impact upon Interpreting the Results
4.3. Geographical Distribution of Viruses in Finland
4.4. Brief Comparison with Other Virome Studies
4.5. Viruses Which Have Pathogenic Associations in Vertebrates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
| Collection No. | Region, Municipality Date | Genome Type | Virus Family/ Taxon | Proposed or Established Name |
|---|---|---|---|---|
| FI 432 | KH, Hattula | –ssRNA | Chuviridae | Hattula chuvirus |
| 27 May 2015 | −ssRNA | Rhabdoviridae | Hattula rhabdovirus | |
| dsRNA | Partitiviridae | Hattula partiti-like virus | ||
| dsRNA | Totiviridae | Hattula totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 2 | ||
| dsRNA | Totiviridae | Hattula totivirus 3 | ||
| FI 437 | KH, Hämeenlinna | −ssRNA | Phasmaviridae | Hameenlinna orthophasmavirus 1 |
| 2 June 2015 | −ssRNA | Phasmaviridae | Hameenlinna orthophasmavirus 2 | |
| −ssRNA | Phenuiviridae | Kalajoki phenui-like virus 2 | ||
| dsRNA | Partitiviridae | Hameenlinna partiti-like virus | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| FI 441, FI 442 | KH, Hämeenlinna | +ssRNA | Flaviviridae | Hameenlinna flavivirus |
| 2 June 2015 | −ssRNA | Phenuiviridae | Hameenlinna phasivirus | |
| dsRNA | Totiviridae | Hameenlinna toti-like virus | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 2 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 3 | ||
| FI 474 | Po, Vaasa | +ssRNA | Iflaviridae | Hanko iflavirus 1 |
| 14 June 2015 | dsRNA | Partitiviridae | Hameenlinna partiti-like virus | |
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| FI 487 | Po, Vaasa | dsRNA | Partitiviridae | Vaasa deltapartitivirus |
| 16 June 2015 | dsRNA | Partitiviridae | Vaasa partiti-like virus | |
| dsRNA | Totiviridae | Vaasa toti-like virus | ||
| dsRNA | Totiviridae | Vaasa totivirus | ||
| FI 500 | Po, Pedersöre | +ssRNA | Iflaviridae | Pedersore iflavirus |
| 17 June 2015 | dsRNA | Totiviridae | Hameenlinna totivirus 1 | |
| dsRNA | Totiviridae | Hattula totivirus 2 | ||
| +ssRNA | Virgaviridae | Pedersore virga-like virus | ||
| FI 505 | PP, Kalajoki | −ssRNA | Chuviridae | Hattula chuvirus |
| 17 June 2015 | −ssRNA | Phenuiviridae | Kalajoki phenui-like virus 1 | |
| −ssRNA | Phenuiviridae | Kalajoki phenui-like virus 2 | ||
| −ssRNA | Qinviridae | Kalajoki qinvirus | ||
| dsRNA | Partitiviridae | Kalajoki alphapartitivirus | ||
| dsRNA | Partitiviridae | Kalajoki betapartitivirus | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 2 | ||
| FI 513 | KP, Lestijärvi | +ssRNA | Flaviviridae | Lestijarvi flavi-like virus |
| 18 June 2015 | +ssRNA | Iflaviridae | Pedersore iflavirus | |
| −ssRNA | Aliusviridae | Lestijarvi obscuruvirus | ||
| −ssRNA | Chuviridae | Hattula chuvirus | ||
| −ssRNA | Phasmaviridae | Lestijarvi orthophasmavirus 1 | ||
| −ssRNA | Phasmaviridae | Lestijarvi orthophasmavirus 2 | ||
| dsRNA | Chrysoviridae | Lestijarvi alphachrysovirus | ||
| dsRNA | Partitiviridae | Lestijarvi partiti-like virus | ||
| dsRNA | Totiviridae | Lestijarvi totivirus | ||
| FI 520 | KS, Karstula | −ssRNA | Chuviridae | Hattula chuvirus |
| 18 June 2015 | −ssRNA | Rhabdoviridae | Hattula rhabdovirus | |
| dsRNA | Totiviridae | Karstula totivirus | ||
| FI 525 | Pi, Pälkäne | −ssRNA | Phasmaviridae | Hameenlinna orthophasmavirus 1 |
| 19 June 2015 | −ssRNA | Phenuiviridae | Kalajoki phenui-like virus 2 | |
| −ssRNA | Phenuiviridae | Palkane phenui-like virus 1 | ||
| −ssRNA | Phenuiviridae | Palkane phenui-like virus 2 | ||
| −ssRNA | Qinviridae | Kalajoki qinvirus | ||
| −ssRNA | Qinviridae | Palkane qinvirus | ||
| dsRNA | Botybirnavirus | Palkane botybirna-like virus | ||
| dsRNA | Partitiviridae | Kalajoki alphapartitivirus | ||
| dsRNA | Partitiviridae | Kalajoki betapartitivirus | ||
| dsRNA | Partitiviridae | Palkane alphapartitivirus 1 | ||
| dsRNA | Partitiviridae | Palkane alphapartitivirus 2 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 2 | ||
| dsRNA | Totiviridae | Palkane toti-like virus | ||
| dsRNA | Totiviridae | Palkane totivirus | ||
| FI 531 | EK, Imatra | dsRNA | Totiviridae | Hameenlinna totivirus 1 |
| 23 June 2015 | dsRNA | Totiviridae | Hattula totivirus 2 | |
| dsRNA | Totiviridae | Palkane toti-like virus | ||
| FI 532, FI 537 | VS, Ilomantsi | +ssRNA | Iflaviridae | Hanko iflavirus 1 |
| 25–26 June 2015 | +ssRNA | Iflaviridae | Mekrijarvi iflavirus | |
| +ssRNA | Negevirus | Mekrijarvi negevirus | ||
| +ssRNA | Solemoviridae | Ilomantsi sobemovirus | ||
| +ssRNA | Togaviridae | Sindbis virus | ||
| −ssRNA | Phenuiviridae | Hameenlinna phasivirus | ||
| −ssRNA | Phenuiviridae | Ilomantsi phenui-like virus | ||
| −ssRNA | Phenuiviridae | Kalajoki phenui-like virus 1 | ||
| −ssRNA | Phenuiviridae | Kalajoki phenui-like virus 2 | ||
| −ssRNA | Qinviridae | Kalajoki qinvirus | ||
| −ssRNA | Qinviridae | Palkane qinvirus | ||
| −ssRNA | Qinviridae | Ilomantsi qinvirus | ||
| −ssRNA | Rhabdoviridae | Hattula rhabdovirus | ||
| dsRNA | Botybirnavirus | Palkane botybirna-like virus | ||
| dsRNA | Partitiviridae | Ilomantsi deltapartitivirus | ||
| dsRNA | Partitiviridae | Ilomantsi partiti-like virus 2 | ||
| dsRNA | Partitiviridae | Kalajoki alphapartitivirus | ||
| dsRNA | Partitiviridae | Lestijarvi partiti-like virus | ||
| dsRNA | Partitiviridae | Vaasa deltapartitivirus | ||
| dsRNA | Partitiviridae | Hameenlinna partiti-like virus | ||
| dsRNA | Sedoreoviridae | Ilomantsi reovirus 1 | ||
| dsRNA | Sedoreoviridae | Ilomantsi reovirus 2 | ||
| dsRNA | Sedoreoviridae | Ilomantsi reovirus 3 | ||
| dsRNA | Totiviridae | Hameenlinna toti-like virus | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 3 | ||
| dsRNA | Totiviridae | Hattula totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 2 | ||
| dsRNA | Totiviridae | Hattula totivirus 3 | ||
| dsRNA | Totiviridae | Ilomantsi toti-like virus 1 | ||
| dsRNA | Totiviridae | Ilomantsi toti-like virus 2 | ||
| dsRNA | Totiviridae | Ilomantsi toti-like virus 3 | ||
| dsRNA | Totiviridae | Ilomantsi totivirus 1 | ||
| dsRNA | Totiviridae | Ilomantsi totivirus 2 | ||
| dsRNA | Totiviridae | Ilomantsi totivirus 3 | ||
| dsRNA | Totiviridae | Lestijarvi totivirus | ||
| dsRNA | Totiviridae | Palkane toti-like virus | ||
| dsRNA | Totiviridae | Vaasa toti-like virus | ||
| dsRNA | Totiviridae | Vaasa totivirus | ||
| FI 538 | VS, Ilomantsi | −ssRNA | Phenuiviridae | Hameenlinna phasivirus |
| 26 June 2015 | dsRNA | Partitiviridae | Ilomantsi partiti-like virus 1 | |
| dsRNA | Totiviridae | Hameenlinna totivirus 3 | ||
| FI 540 | VS, Ilomantsi | −ssRNA | Phenuiviridae | Kalajoki phenui-like virus 1 |
| 26 June 2015 | dsRNA | Totiviridae | Hameenlinna totivirus 1 | |
| FI 550 | VS, Joensuu | +ssRNA | Solemoviridae | Joensuu sobemovirus |
| 7 June 2015 | −ssRNA | Chuviridae | Hattula chuvirus | |
| −ssRNA | Phenuiviridae | Hameenlinna phasivirus | ||
| −ssRNA | Rhabdoviridae | Hattula rhabdovirus | ||
| −ssRNA | Xinmoviridae | Joensuu anphevirus | ||
| dsRNA | Partitiviridae | Ilomantsi partiti-like virus 2 | ||
| dsRNA | Partitiviridae | Hattula partiti-like virus | ||
| dsRNA | Sedoreoviridae | Ilomantsi reovirus 2 | ||
| dsRNA | Sedoreoviridae | Ilomantsi reovirus 4 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 2 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 3 | ||
| dsRNA | Totiviridae | Hattula totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 2 | ||
| dsRNA | Totiviridae | Ilomantsi toti-like virus 2 | ||
| dsRNA | Totiviridae | Ilomantsi totivirus 2 | ||
| dsRNA | Totiviridae | Palkane toti-like virus | ||
| FI 551 | VS, Ilomantsi | +ssRNA | Iflaviridae | Hanko iflavirus 1 |
| 27 June 2015 | −ssRNA | Phenuiviridae | Hameenlinna phasivirus | |
| dsRNA | Sedoreoviridae | Ilomantsi reovirus 1 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 2 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 3 | ||
| FI 566 | PP, Kiiminki 3 July 2015 | dsRNA | Partitiviridae | Ilomantsi deltapartitivirus |
| FI 571 | L, Kittilä | dsRNA | Partitiviridae | Ilomantsi partiti-like virus 1 |
| 3 July 2015 | dsRNA | Totiviridae | Hameenlinna toti-like virus | |
| dsRNA | Totiviridae | Hameenlinna totivirus 2 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 3 | ||
| FI 575, FI 730 | L, Enontekiö 3–4 July 2015 | +ssRNA | Iflaviridae | Hanko iflavirus 1 |
| FI 575, FI 582, FI 728 | L, Enontekiö | dsRNA | Partitiviridae | Hattula partiti-like virus |
| 2 and 4 July 2015 | dsRNA | Totiviridae | Enontekio totivirus 5 | |
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| FI 607 | L, Utsjoki | +ssRNA | Negevirus | Cordoba virus |
| 7 July 2015 | +ssRNA | Negevirus | Utsjoki negevirus 1 | |
| +ssRNA | Negevirus | Utsjoki negevirus 2 | ||
| +ssRNA | Negevirus | Utsjoki negevirus 3 | ||
| −ssRNA | Chuviridae | Hattula chuvirus | ||
| dsRNA | Totiviridae | Hattula totivirus 1 | ||
| FI 618 | PP, Kuusamo | +ssRNA | Iflaviridae | Hanko iflavirus 1 |
| 9 July 2015 | −ssRNA | Chuviridae | Hattula chuvirus | |
| dsRNA | Chrysoviridae | Lestijarvi alphachrysovirus | ||
| FI 618, FI 620 | PP, Kuusamo | dsRNA | Partitiviridae | Ilomantsi partiti-like virus 1 |
| 9 July 2015 | dsRNA | Totiviridae | Kuusamo totivirus 1 | |
| dsRNA | Totiviridae | Kuusamo totivirus 2 | ||
| FI 620 | PP, Kuusamo | +ssRNA | Negevirus | Cordoba virus |
| 9 July 2015 | +ssRNA | Negevirus | Cordoba virus | |
| +ssRNA | Negevirus | Utsjoki negevirus 1 | ||
| dsRNA | Partitiviridae | Hattula partiti-like virus | ||
| dsRNA | Totiviridae | Enontekio totivirus 5 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| FI 641 | PP, Kuusamo | +ssRNA | Negevirus | Cordoba virus |
| 18 July 2015 | +ssRNA | Negevirus | Dezidougou virus | |
| +ssRNA | Negevirus | Utsjoki negevirus 1 | ||
| +ssRNA | Solemoviridae | Joensuu sobemovirus | ||
| −ssRNA | Phasmaviridae | Kuusamo orthophasmavirus 1 | ||
| −ssRNA | Phasmaviridae | Kuusamo orthophasmavirus 2 | ||
| −ssRNA | Phasmaviridae | Kuusamo orthophasmavirus 3 | ||
| −ssRNA | Phasmaviridae | Kuusamo orthophasmavirus 4 | ||
| −ssRNA | Xinmoviridae | Joensuu anphevirus | ||
| dsRNA | Partitiviridae | Kuusamo partiti-like virus | ||
| dsRNA | Partitiviridae | Kuusamo alphapartitivirus | ||
| dsRNA | Partitiviridae | Hattula partiti-like virus | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Ilomantsi totivirus 1 | ||
| dsRNA | Totiviridae | Kuusamo toti-like virus | ||
| dsRNA | Totiviridae | Lestijarvi totivirus | ||
| FI 642, FI 648 | L, Salla 19 July 2015 | +ssRNA | Iflaviridae | Hanko iflavirus 1 |
| FI 649 | L, Inari | +ssRNA | Flaviviridae | Inari jingmenvirus |
| 22 July 2015 | +ssRNA | Negevirus | Utsjoki negevirus 1 | |
| +ssRNA | Negevirus | Utsjoki negevirus 2 | ||
| +ssRNA | Permutotetraviridae | Inari permutotetravirus | ||
| −ssRNA | Rhabdoviridae | Hattula rhabdovirus | ||
| −ssRNA | Rhabdoviridae | Inari rhabdovirus | ||
| −ssRNA | Rhabdoviridae | Ohlsdorf virus | ||
| dsRNA | Partitiviridae | Ilomantsi partiti-like virus 1 | ||
| dsRNA | Partitiviridae | Hattula partiti-like virus | ||
| dsRNA | Partitiviridae | Inari deltapartitivirus | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 3 | ||
| dsRNA | Totiviridae | Ilomantsi toti-like virus 2 | ||
| dsRNA | Totiviridae | Inari toti-like virus | ||
| dsRNA | Totiviridae | Inari totivirus 1 | ||
| dsRNA | Totiviridae | Inari totivirus 2 | ||
| dsRNA | Totiviridae | Palkane toti-like virus | ||
| dsRNA | Totiviridae | Vaasa toti-like virus | ||
| FI 649, FI 654, FI 655 | L, Inari | +ssRNA | Permutotetraviridae | Inari permutotetravirus |
| 22–23 July 2015 | +ssRNA | Solemoviridae | Ilomantsi sobemovirus | |
| −ssRNA | Rhabdoviridae | Ohlsdorf virus | ||
| dsRNA | Partitiviridae | Vaasa deltapartitivirus | ||
| dsRNA | Totiviridae | Ilomantsi toti-like virus 2 | ||
| dsRNA | Totiviridae | Inari totivirus 1 | ||
| dsRNA | Totiviridae | Vaasa toti-like virus | ||
| FI 652, FI 728 | L, Inari/Enontekiö | dsRNA | Partitiviridae | Enontekio partiti-like virus |
| 2 July 2015 | dsRNA | Totiviridae | Enontekio totivirus 3 | |
| dsRNA | Totiviridae | Enontekio totivirus 4 | ||
| dsRNA | Totiviridae | Hattula totivirus 3 | ||
| FI 654, FI 655 | L, Inari | +ssRNA | Solemoviridae | Joensuu sobemovirus |
| 23 July 2015 | dsRNA | Totiviridae | Inari totivirus 2 | |
| dsRNA | Totiviridae | Lestijarvi totivirus | ||
| FI 663 | L, Utsjoki | +ssRNA | Virgaviridae | Pedersore virga-like virus |
| 24 July 2015 | dsRNA | Chrysoviridae | Lestijarvi alphachrysovirus | |
| dsRNA | Partitiviridae | Hameenlinna partiti-like virus | ||
| dsRNA | Sedoreoviridae | Ilomantsi reovirus 1 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 1 | ||
| dsRNA | Totiviridae | Inari totivirus 2 | ||
| dsRNA | Totiviridae | Utsjoki toti-like virus | ||
| FI 671 | L, Enontekiö | +ssRNA | Iflaviridae | Enontekio iflavirus |
| 26 July 2015 | +ssRNA | Negevirus | Cordoba virus | |
| +ssRNA | Negevirus | Utsjoki negevirus 1 | ||
| +ssRNA | Negevirus | Utsjoki negevirus 2 | ||
| +ssRNA | Negevirus | Utsjoki negevirus 3 | ||
| +ssRNA | Solemoviridae | Enontekio sobemovirus | ||
| −ssRNA | Chuviridae | Hattula chuvirus | ||
| −ssRNA | Rhabdoviridae | Enontekio rhabdovirus | ||
| dsRNA | Chrysoviridae | Enontekio alphachrysovirus | ||
| dsRNA | Partitiviridae | Ilomantsi partiti-like virus 1 | ||
| dsRNA | Totiviridae | Enontekio totivirus 2 | ||
| dsRNA | Totiviridae | Hattula totivirus 3 | ||
| dsRNA | Totiviridae | Hattula totivirus 3 | ||
| FI 674 | L, Enontekiö | +ssRNA | Flaviviridae | Kilpisjarvi flavivirus |
| 28 July 2015 | +ssRNA | Iflaviridae | Enontekio iflavirus | |
| +ssRNA | Negevirus | Utsjoki negevirus 1 | ||
| +ssRNA | Permutotetraviridae | Inari permutotetravirus | ||
| +ssRNA | Quenyavirus | Enontekio quenyavirus | ||
| +ssRNA | Solemoviridae | Enontekio sobemovirus | ||
| +ssRNA | Virgaviridae | Enontekio virga-like virus 1 | ||
| +ssRNA | Virgaviridae | Enontekio virga-like virus 2 | ||
| +ssRNA | Virgaviridae | Pedersore virga-like virus | ||
| −ssRNA | Aspiviridae | Kilpisjarvi aspivirus | ||
| −ssRNA | Phenuiviridae | Enontekio phenui-like virus 1 | ||
| −ssRNA | Phenuiviridae | Enontekio phenui-like virus 3 | ||
| −ssRNA | Phenuiviridae | Enontekio phenui-like virus 5 | ||
| −ssRNA | Rhabdoviridae | Enontekio merhavirus | ||
| −ssRNA | Rhabdoviridae | Enontekio ohlsrhavirus | ||
| −ssRNA | Xinmoviridae | Enontekio anphevirus 1 | ||
| −ssRNA | Xinmoviridae | Enontekio anphevirus 2 | ||
| −ssRNA | Yueviridae | Enontekio yuevirus | ||
| dsRNA | Partitiviridae | Enontekio alphapartitivirus 1 | ||
| dsRNA | Partitiviridae | Enontekio alphapartitivirus 2 | ||
| dsRNA | Partitiviridae | Enontekio betapartitivirus 1 | ||
| dsRNA | Partitiviridae | Enontekio betapartitivirus 2 | ||
| dsRNA | Partitiviridae | Ilomantsi partiti-like virus 1 | ||
| dsRNA | Partitiviridae | Kuusamo alphapartitivirus | ||
| dsRNA | Spinareoviridae | Enontekio reovirus | ||
| dsRNA | Totiviridae | Enontekio toti-like virus 1 | ||
| dsRNA | Totiviridae | Enontekio toti-like virus 2 | ||
| dsRNA | Totiviridae | Enontekio toti-like virus 3 | ||
| dsRNA | Totiviridae | Enontekio toti-like virus 4 | ||
| dsRNA | Totiviridae | Enontekio totivirus 1 | ||
| dsRNA | Totiviridae | Enontekio totivirus 2 | ||
| dsRNA | Totiviridae | Enontekio totivirus 6 | ||
| dsRNA | Totiviridae | Enontekio totivirus 7 | ||
| dsRNA | Totiviridae | Palkane toti-like virus | ||
| FI 675 | L, Enontekiö | +ssRNA | Flaviviridae | Inari jingmenvirus |
| 28 July 2015 | −ssRNA | Rhabdoviridae | Enontekio ohlsrhavirus | |
| FI 701 | PP, Kuusamo | +ssRNA | Negevirus | Cordoba virus |
| 23 August 2015 | +ssRNA | Negevirus | Utsjoki negevirus 1 | |
| +ssRNA | Negevirus | Utsjoki negevirus 2 | ||
| +ssRNA | Permutotetraviridae | Inari permutotetravirus | ||
| FI 976 | EK, Joutseno | +ssRNA | Virgaviridae | Pedersore virga-like virus |
| 4 July 2017 | −ssRNA | Chuviridae | Hattula chuvirus | |
| −ssRNA | Phasmaviridae | Hameenlinna orthophasmavirus 1 | ||
| −ssRNA | Phasmaviridae | Hameenlinna orthophasmavirus 2 | ||
| −ssRNA | Phasmaviridae | Kuusamo orthophasmavirus 4 | ||
| −ssRNA | Phenuiviridae | Enontekio phenui-like virus 2 | ||
| −ssRNA | Phenuiviridae | Palkane phenui-like virus 2 | ||
| −ssRNA | Rhabdoviridae | Joutseno rhabdovirus 1 | ||
| −ssRNA | Rhabdoviridae | Joutseno rhabdovirus 2 | ||
| dsRNA | Partitiviridae | Hattula partiti-like virus | ||
| dsRNA | Sedoreoviridae | Ilomantsi reovirus 2 | ||
| dsRNA | Totiviridae | Hameenlinna totivirus 1 | ||
| dsRNA | Totiviridae | Hattula totivirus 2 | ||
| dsRNA | Totiviridae | Joutseno totivirus | ||
| dsRNA | Totiviridae | Palkane totivirus | ||
| FI 988 | VS, Kustavi | +ssRNA | Solemoviridae | Evros sobemo-like virus |
| 11 July 2017 | −ssRNA | Chuviridae | Kustavi chuvirus 1 | |
| −ssRNA | Chuviridae | Kustavi chuvirus 2 | ||
| dsRNA | Totiviridae | Kustavi toti-like virus | ||
| FI 1009, FI 1010, FI 1011 | U, Hanko | +ssRNA | Flaviviridae | Hanko virus |
| 22–23 August 2017 | +ssRNA | Iflaviridae | Hanko iflavirus 2 | |
| +ssRNA | Iflaviridae | Pedersore iflavirus | ||
| +ssRNA | Picornaviridae | Hanko picorna-like virus | ||
| +ssRNA | Quenyavirus | Enontekio quenyavirus | ||
| +ssRNA | Solemoviridae | Evros sobemo-like virus | ||
| +ssRNA | Solemoviridae | Hanko sobemovirus | ||
| +ssRNA | Virgaviridae | Pedersore virga-like virus | ||
| −ssRNA | Phenuiviridae | Enontekio phenui-like virus 4 | ||
| −ssRNA | Phenuiviridae | Hanko phenui-like virus 1 | ||
| −ssRNA | Phenuiviridae | Hanko phenui-like virus 2 | ||
| −ssRNA | Phenuiviridae | Hanko phenui-like virus 3 | ||
| −ssRNA | Xinmoviridae | Hanko anphevirus | ||
| −ssRNA | Xinmoviridae | Joensuu anphevirus | ||
| dsRNA | Chrysoviridae | Hanko alphachrysovirus | ||
| dsRNA | Endornaviridae | Hallsjon virus | ||
| dsRNA | Endornaviridae | Tvarminne alphaendornavirus | ||
| dsRNA | Partitiviridae | Hanko alphapartitivirus 1 | ||
| dsRNA | Partitiviridae | Hanko alphapartitivirus 2 | ||
| dsRNA | Partitiviridae | Hanko alphapartitivirus 3 | ||
| dsRNA | Totiviridae | Hanko toti-like virus 1 | ||
| dsRNA | Totiviridae | Hanko toti-like virus 2 | ||
| dsRNA | Totiviridae | Hanko toti-like virus 3 | ||
| dsRNA | Totiviridae | Hanko totivirus 1 | ||
| dsRNA | Totiviridae | Hanko totivirus 2 | ||
| dsRNA | Totiviridae | Hanko totivirus 3 | ||
| dsRNA | Totiviridae | Hanko totivirus 4 | ||
| dsRNA | Totiviridae | Hanko totivirus 5 | ||
| dsRNA | Totiviridae | Hanko totivirus 6 | ||
| dsRNA | Totiviridae | Hanko totivirus 7 | ||
| dsRNA | Totiviridae | Hanko totivirus 8 | ||
| dsRNA | Totiviridae | Hanko totivirus 9 | ||
| dsRNA | Totiviridae | Hanko totivirus 10 | ||
| dsRNA | Totiviridae | Hattula totivirus 3 | ||
| dsRNA | Totiviridae | Inari toti-like virus | ||
| dsRNA | Totiviridae | Kustavi toti-like virus | ||
| FI 1015 | VS, Kustavi | +ssRNA | Negevirus | Kustavi negevirus |
| 24 August 2017 | +ssRNA | Picornaviridae | Jotan virus | |
| +ssRNA | Solemoviridae | Evros sobemo-like virus | ||
| dsRNA | Totiviridae | Hanko totivirus 3 | ||
| dsRNA | Totiviridae | Hanko totivirus 4 | ||
| dsRNA | Totiviridae | Hanko totivirus 5 | ||
| dsRNA | Totiviridae | Kustavi toti-like virus |
References
- Sane, J.; Kurkela, S.; Putkuri, N.; Huhtamo, E.; Vaheri, A.; Vapalahti, O. Complete coding sequence and molecular epidemiological analysis of Sindbis virus isolates from mosquitoes and humans, Finland. J. Gen. Virol. 2012, 93, 1984–1990. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, E.M.; Suvanto, M.T.; Uusitalo, R.; Faolotto, G.; Smura, T.; Sane, J.; Vapalahti, O.; Huhtamo, E. Sindbis virus strains of divergent origin isolated from humans and mosquitoes during a recent outbreak in Finland. Vector Borne Zoonotic Dis. 2020, 20, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Brummer-Korvenkontio, M.; Saikku, P.; Korhonen, P.; Reunala, T. The Inkoo virus, a member of the California encephalitis group in Finland. Scand. J. Clin. Lab. Invest. 1969, 23 (Suppl. 108), 159. [Google Scholar]
- Brummer-Korvenkontio, M.; Saikku, P.; Korhonen, P.; Ulmanen, I.; Reunala, T.; Karvonen, J. Arboviruses in Finland. IV. Isolation and characterization of Inkoo virus, a Finnish representative of the California group. Am. J. Trop. Med. Hyg. 1973, 22, 404–413. [Google Scholar] [CrossRef]
- Putkuri, N.; Kurkela, S.; Levanov, L.; Huhtamo, E.; Vaheri, A.; Sironen, T.; Vapalahti, O. Isolation and characterization of a California encephalitis serogroup orthobunyavirus from Finnish mosquitoes. Infect. Genet. Evol. 2014, 22, 164–173. [Google Scholar] [CrossRef]
- Putkuri, N.; Kantele, A.; Levanov, L.; Kivisto, I.; Brummer-Korvenkontio, M.; Vaheri, A.; Vapalahti, O. Acute human Inkoo and Chatanga Virus infections, Finland. Emerg Infect. Dis. 2016, 22, 810–817. [Google Scholar] [CrossRef]
- Huhtamo, E.; Putkuri, N.; Kurkela, S.; Manni, T.; Vaheri, A.; Vapalahti, O.; Uzcátegui, N.Y. Characterization of a novel flavivirus from mosquitoes in northern Europe that is related to mosquito-borne flaviviruses of the tropics. J. Virol. 2009, 83, 9532–9540. [Google Scholar] [CrossRef]
- Huhtamo, E.; Moureau, G.; Cook, S.; Julkunen, O.; Putkuri, N.; Kurkela, S.; Uzcátegui, N.Y.; Harbach, R.E.; Gould, E.A.; Vapalahti, O.; et al. Novel insect-specific flavivirus isolated from northern Europe. Virology 2012, 433, 471–478. [Google Scholar] [CrossRef]
- Huhtamo, E.; Cook, S.; Moureau, G.; Uzcátegui, N.Y.; Sironen, T.; Kuivanen, S.; Putkuri, N.; Kurkela, S.; Harbach, R.E.; Firth, A.E.; et al. Novel flaviviruses from mosquitoes: Mosquito-specific evolutionary lineages within the phylogenetic group of mosquito-borne flaviviruses. Virology 2014, 464–465, 320–329. [Google Scholar] [CrossRef]
- Suvanto, M.T.; Truong Nguyen, P.; Uusitalo, R.; Korhonen, E.M.; Faolotto, G.; Vapalahti, O.; Huhtamo, E.; Smura, T. A novel negevirus isolated from Aedes vexans mosquitoes in Finland. Arch. Virol. 2020, 165, 2989–2992. [Google Scholar] [CrossRef]
- Culverwell, C.L.; Uusitalo, R.J.; Korhonen, E.M.; Vapalahti, O.P.; Huhtamo, E.; Harbach, R.E. The mosquitoes of Finland: Updated distributions and bionomics. Med. Vet. Entomol. 2021, 35, 1–29. [Google Scholar] [CrossRef]
- Utrio, P. Geographic distribution of mosquitoes (Diptera, Culicidae) in eastern Fennoscandia. Neotrop. Entomol. 1979, 59, 105–123. [Google Scholar]
- Harbach, R.E. Mosquito Taxonomic Inventory. Available online: http://mosquito-taxonomic-inventory.info (accessed on 7 April 2022).
- ECDC. Guidelines for the Surveillance of Native Mosquitoes in Europe; European Centre for Disease Prevention and Control, Stockholm: Stockholm, Sweden, 2014. [Google Scholar]
- Stojanovich, C.J.; Scott, H.G. Illustrated Key to the Mosquitoes of Fennoscandia: Finland, Sweden, Denmark, Norway; Scott: Portland, OR, USA, 1995; p. 132. [Google Scholar]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.B.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Conceição-Neto, N.; Zeller, M.; Lefrère, H.; De Bruyn, P.; Beller, L.; Deboutte, W.; Yinda, C.K.; Lavigne, R.; Maes, P.; Ranst, M.V.; et al. Modular approach to customise sample preparation procedures for viral metagenomics: A reproducible protocol for virome analysis. Sci. Rep. 2015, 5, 16532. [Google Scholar] [CrossRef]
- Plyusnin, I.; Kant, R.; Jääskelainen, A.J.; Sironen, T.; Holm, L.; Vapalahti, O.; Smura, T. Novel NGS pipeline for virus discovery from a wide spectrum of hosts and sample types. Virus Evol. 2020, 6, veaa091. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, D.; Luo, R.; Liu, C.-M.; Leung, C.-M.; Ting, H.-F.; Sadakane, K.; Yamashita, H.; Lam, T.-W. MEGAHIT v1.0: A fast and scalable metagenome assembler driven by advanced methodologies and community practices. Methods 2016, 102, 3–11. [Google Scholar] [CrossRef]
- Noguchi, H.; Taniguchi, T.; Itoh, T. MetaGeneAnnotator: Detecting species-specific patterns of ribosomal binding site for precise gene prediction in anonymous prokaryotic and phage genomes. DNA Res. 2008, 15, 387–396. [Google Scholar] [CrossRef]
- Stajich, J.E.; Block, D.; Boulez, K.; Brenner, S.E.; Chervitz, S.A.; Dagdigian, C.; Fuellen, G.; Gilbert, J.G.; Korf, I.; Lapp, H.; et al. The Bioperl toolkit: Perl modules for the life sciences. Genome Res. 2002, 12, 1611–1618. [Google Scholar] [CrossRef]
- Somervuo, P.; Holm, L. SANSparallel: Interactive homology search against Uniprot. Nucleic Acids Res. 2015, 43, W24–W29. [Google Scholar] [CrossRef]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. GGTREE: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Pettersson, J.H.; Shi, M.; Eden, J.S.; Holmes, E.C.; Hesson, J.C. Meta-transcriptomic comparison of the RNA viromes of the mosquito vectors Culex pipiens and Culex torrentium in Northern Europe. Viruses 2019, 11, 1033. [Google Scholar] [CrossRef]
- Shahhosseini, N.; Lühken, R.; Jöst, H.; Jansen, S.; Börstler, J.; Rieger, T.; Krüger, A.; Yadouleton, A.; de Mendonça Campos, R.; Cirne-Santos, C.C.; et al. Detection and characterization of a novel rhabdovirus in Aedes cantans mosquitoes and evidence for a mosquito-associated new genus in the family Rhabdoviridae. Infect. Genet. Evol. 2017, 55, 260–268. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, Á.; Pál, J.; Kapusinszky, B.; Delwart, E.; Pankovics, P. Detection and genome analysis of a novel (dima)rhabdovirus (Riverside virus) from Ochlerotatus sp. mosquitoes in Central Europe. Infect. Genet. Evol. 2016, 39, 336–341. [Google Scholar] [CrossRef]
- Sadeghi, M.; Altan, E.; Deng, X.; Barker, C.M.; Fang, Y.; Coffey, L.L.; Delwart, E. Virome of > 12 thousand Culex mosquitoes from throughout California. Virology 2018, 523, 74–88. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, J.H.; Shi, M.; Bohlin, J.; Eldholm, V.; Brynildsrud, O.B.; Paulsen, K.M.; Andreassen, Å.; Holmes, E.C. Characterizing the virome of Ixodes ricinus ticks from northern Europe. Sci. Rep. 2017, 7, 10870. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Neville, P.; Nicholson, J.; Eden, J.S.; Imrie, A.; Holmes, E.C. High-resolution metatranscriptomics reveals the ecological dynamics of mosquito-associated RNA viruses in Western Australia. J. Virol. 2017, 91, e00680-17. [Google Scholar] [CrossRef] [PubMed]
- Faizah, A.N.; Kobayashi, D.; Isawa, H.; Amoa-Bosompem, M.; Murota, K.; Higa, Y.; Futami, K.; Shimada, S.; Kim, K.S.; Itokawa, K.; et al. Deciphering the virome of Culex vishnui subgroup mosquitoes, the major vectors of Japanese Encephalitis, in Japan. Viruses 2020, 12, 264. [Google Scholar] [CrossRef]
- Valverde, R.A.; Khalifa, M.E.; Okada, R.; Fukuhara, T.; Sabanadzovic, S.; Ictv Report, C. ICTV Virus Taxonomy Profile: Endornaviridae. J. Gen. Virol. 2019, 100, 1204–1205. [Google Scholar] [CrossRef]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef]
- Akiner, M.M.; Ozturk, M.; Baser, A.B.; Gunay, F.; Hacioglu, S.; Brinkmann, A.; Emanet, N.; Alten, B.; Ozkul, A.; Nitsche, A.; et al. Arboviral screening of invasive Aedes species in northeastern Turkey: West Nile virus circulation and detection of insect-only viruses. PLoS Negl. Trop. Dis. 2019, 13, e0007334. [Google Scholar] [CrossRef]
- Supriyono, R.K.; Torii, S.; Shimoda, H.; Ishijima, K.; Yonemitsu, K.; Minami, S.; Kuroda, Y.; Tatemoto, K.; Tran, N.T.B.; Takano, A.; et al. Mosquito-borne viruses, insect-specific flaviviruses (family Flaviviridae, genus Flavivirus), Banna virus (family Reoviridae, genus Seadornavirus), Bogor virus (unassigned member of family Permutotetraviridae), and alphamesoniviruses 2 and 3 (family Mesoniviridae, genus Alphamesonivirus) isolated from Indonesian mosquitoes. J. Vet. Med. Sci. 2020, 82, 1030–1041. [Google Scholar] [CrossRef]
- He, X.; Yin, Q.; Zhou, L.; Meng, L.; Hu, W.; Li, F.; Li, Y.; Han, K.; Zhang, S.; Fu, S.; et al. Metagenomic sequencing reveals viral abundance and diversity in mosquitoes from the Shaanxi-Gansu-Ningxia region, China. PLoS Negl. Trop. Dis. 2021, 15, e0009381. [Google Scholar] [CrossRef]
- Valles, S.M.; Chen, Y.; Firth, A.E.; Guerin, D.M.A.; Hashimoto, Y.; Herrero, S.; de Miranda, J.R.; Ryabov, E.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Iflaviridae. J Gen. Virol. 2017, 98, 527–528. [Google Scholar] [CrossRef]
- Atoni, E.; Wang, Y.; Karungu, S.; Waruhiu, C.; Zohaib, A.; Obanda, V.; Agwanda, B.; Mutua, M.; Xia, H.; Yuan, Z. Metagenomic virome analysis of Culex mosquitoes from Kenya and China. Viruses 2018, 10, 30. [Google Scholar] [CrossRef]
- Vasilakis, N.; Forrester, N.L.; Palacios, G.; Nasar, F.; Savji, N.; Rossi, S.L.; Guzman, H.; Wood, T.G.; Popov, V.; Gorchakov, R.; et al. Negevirus: A proposed new taxon of insect-specific viruses with wide geographic distribution. J. Virol. 2013, 87, 2475–2488. [Google Scholar] [CrossRef]
- Pringle, F.M.; Gordon, K.H.; Hanzlik, T.N.; Kalmakoff, J.; Scotti, P.D.; Ward, V.K. A novel capsid expression strategy for Thosea asigna virus (Tetraviridae). J Gen. Virol. 1999, 80 Pt 7, 1855–1863. [Google Scholar] [CrossRef]
- Zeddam, J.-L.; Gordon, K.H.J.; Lauber, C.; Alves, C.A.F.; Luke, B.T.; Hanzlik, T.N.; Ward, V.K.; Gorbalenya, A.E. Euprosterna elaeasa virus genome sequence and evolution of the Tetraviridae family: Emergence of bipartite genomes and conservation of the VPg signal with the dsRNA Birnaviridae family. Virology 2010, 397, 145–154. [Google Scholar] [CrossRef]
- Ruiz-Padilla, A.; Rodríguez-Romero, J.; Gómez-Cid, I.; Pacifico, D.; Ayllón, M.A. Novel mycoviruses discovered in the mycovirome of a necrotrophic fungus. Mbio 2021, 12, e03705-20. [Google Scholar] [CrossRef]
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV Virus Taxonomy Profile: Picornaviridae. J Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef]
- Cholleti, H.; Hayer, J.; Fafetine, J.; Berg, M.; Blomström, A.-L. Genetic characterization of a novel picorna-like virus in Culex spp. mosquitoes from Mozambique. Virol. J. 2018, 15, 71. [Google Scholar] [CrossRef]
- Harvey, E.; Rose, K.; Eden, J.-S.; Lawrence, A.; Doggett, S.L.; Holmes, E.C. Identification of diverse arthropod associated viruses in native Australian fleas. Virology 2019, 535, 189–199. [Google Scholar] [CrossRef]
- Obbard, D.J.; Shi, M.; Roberts, K.E.; Longdon, B.; Dennis, A.B. A new lineage of segmented RNA viruses infecting animals. Virus Evol. 2020, 6, vez061. [Google Scholar] [CrossRef]
- Sõmera, M.; Fargette, D.; Hébrard, E.; Sarmiento, C.; Ictv Report, C. ICTV Virus Taxonomy Profile: Solemoviridae 2021. J. Gen. Virol. 2021, 102, 001707. [Google Scholar] [CrossRef]
- Chen, R.; Mukhopadhyay, S.; Merits, A.; Bolling, B.; Nasar, F.; Coffey, L.L.; Powers, A.; Weaver, S.C.; Consortium, I.R. ICTV Virus Taxonomy Profile: Togaviridae. J. Gen. Virol. 2018, 99, 761–762. [Google Scholar] [CrossRef]
- Adams, M.J.; Adkins, S.; Bragard, C.; Gilmer, D.; Li, D.; MacFarlane, S.A.; Wong, S.M.; Melcher, U.; Ratti, C.; Ryu, K.H.; et al. ICTV Virus Taxonomy Profile: Virgaviridae. J. Gen. Virol. 2017, 98, 1999–2000. [Google Scholar] [CrossRef]
- Charon, J.; Murray, S.; Holmes, E.C. Revealing RNA virus diversity and evolution in unicellular algae transcriptomes. Virus Evol. 2021, 7, veab070. [Google Scholar] [CrossRef]
- García, M.L.; Bó, E.D.; da Graça, J.V.; Gago-Zachert, S.; Hammond, J.; Moreno, P.; Natsuaki, T.; Pallás, V.; Navarro, J.A.; Reyes, C.A.; et al. ICTV Virus Taxonomy Profile: Ophioviridae. J. Gen. Virol. 2017, 98, 1161–1162. [Google Scholar] [CrossRef]
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, É.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: Update 2019. Arch. Virol. 2019, 164, 1949–1965. [Google Scholar] [CrossRef]
- De Lara Pinto, A.Z.; Santos de Carvalho, M.; de Melo, F.L.; Ribeiro, A.L.M.; Ribeiro, B.M.; Slhessarenko, R.D. Novel viruses in salivary glands of mosquitoes from sylvatic Cerrado, Midwestern Brazil. PLoS ONE 2017, 12, e0187429. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Adkins, S.; Agwanda, B.R.; Al Kubrusli, R.; Alkhovsky, S.V.; Amarasinghe, G.K.; Avšič-Županc, T.; Ayllón, M.A.; Bahl, J.; Balkema-Buschmann, A.; et al. 2021 Taxonomic update of phylum Negarnaviricota (Riboviria: Orthornavirae), including the large orders Bunyavirales and Mononegavirales. Arch. Virol. 2021, 166, 3513–3566. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Morais, P.; Maia, C.; de Sousa, C.B.; de Almeida, A.P.G.; Parreira, R. A diverse assemblage of RNA and DNA viruses found in mosquitoes collected in southern Portugal. Virus Res. 2019, 274, 197769. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Pang, R.; Cheng, T.; Xue, L.; Zeng, H.; Lei, T.; Chen, M.; Wu, S.; Ding, Y.; Zhang, J.; et al. Abundant and diverse RNA viruses in insects revealed by RNA-Seq analysis: Ecological and evolutionary emplications. mSystems 2020, 5, e00039-20. [Google Scholar] [CrossRef] [PubMed]
- Chiapello, M.; Bosco, L.; Ciuffo, M.; Ottati, S.; Salem, N.; Rosa, C.; Tavella, L.; Turina, M.; Pfeiffer, J.K. Complexity and local specificity of the virome associated with tospovirus-transmitting Thrips species. J. Virol. 2021, 95, e00597-21. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; Junglen, S.; et al. Changes to virus taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2019). Arch. Virol. 2019, 164, 2417–2429. [Google Scholar] [CrossRef]
- Fauver, J.R.; Grubaugh, N.D.; Krajacich, B.J.; Weger-Lucarelli, J.; Lakin, S.M.; Fakoli, L.S., 3rd; Bolay, F.K.; Diclaro, J.W., 2nd; Dabire, K.R.; Foy, B.D.; et al. West African Anopheles gambiae mosquitoes harbor a taxonomically diverse virome including new insect-specific flaviviruses, mononegaviruses, and totiviruses. Virology 2016, 498, 288–299. [Google Scholar] [CrossRef]
- Parry, R.; Asgari, S.; Williams, B.R.G. Aedes anphevirus: An insect-specific virus distributed worldwide in Aedes aegypti mosquitoes that has complex interplays with Wolbachia and Dengue Virus infection in cells. J. Virol. 2018, 92, e00224-18. [Google Scholar] [CrossRef]
- Käfer, S.; Paraskevopoulou, S.; Zirkel, F.; Wieseke, N.; Donath, A.; Petersen, M.; Jones, T.C.; Liu, S.; Zhou, X.; Middendorf, M.; et al. Re-assessing the diversity of negative strand RNA viruses in insects. PLoS Pathog. 2019, 15, e1008224. [Google Scholar] [CrossRef]
- Zhai, L.; Yang, M.; Zhang, M.; Hong, N.; Wang, G. Characterization of a Botybirnavirus conferring hypovirulence in the phytopathogenic fungus Botryosphaeria dothidea. Viruses 2019, 11, 266. [Google Scholar] [CrossRef]
- Ghabrial, S.A.; Caston, J.R.; Coutts, R.H.A.; Hillman, B.I.; Jiang, D.; Kim, D.H.; Moriyama, H.; Ictv Report, C. ICTV Virus Taxonomy Profile: Chrysoviridae. J Gen. Virol. 2018, 99, 19–20. [Google Scholar] [CrossRef]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Partitiviridae. J Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef]
- Duan, Y.; Yang, Z.; Bellis, G.; Xie, J.; Li, L. Full genome sequencing of three Sedoreoviridae viruses isolated from Culicoides spp. (Diptera, Ceratopogonidae) in China. Viruses 2022, 14, 971. [Google Scholar] [CrossRef]
- Attoui, H.; Jaafar, F.M.; Belhouchet, M.; Aldrovandi, N.; Tao, S.; Chen, B.; Liang, G.; Tesh, R.B.; de Micco, P.; de Lamballerie, X. Yunnan orbivirus, a new orbivirus species isolated from Culex tritaeniorhynchus mosquitoes in China. J. Gen. Virol. 2005, 86, 3409–3417. [Google Scholar] [CrossRef]
- Attoui, H.; Stirling, J.M.; Munderloh, U.G.; Billoir, F.; Brookes, S.M.; Burroughs, J.N.; de Micco, P.; Mertens, P.P.C.; de Lamballerie, X. Complete sequence characterization of the genome of the St Croix River virus, a new orbivirus isolated from cells of Ixodes scapularis. J. Gen. Virol. 2001, 82, 795–804. [Google Scholar] [CrossRef]
- Murphy, M.D.; Howerth, E.W.; MacLachlan, N.J.; Stallknecht, D.E. Genetic variation among epizootic hemorrhagic disease viruses in the southeastern United States: 1978–2001. Infect. Genet. Evol. 2005, 5, 157–165. [Google Scholar] [CrossRef]
- Liu, H.; Li, M.-H.; Zhai, Y.-G.; Meng, W.-S.; Sun, X.-H.; Cao, Y.-X.; Fu, S.-H.; Wang, H.-Y.; Xu, L.-H.; Tang, Q.; et al. Banna virus, China, 1987–2007. Emerg. Infect. Dis. 2010, 16, 514–517. [Google Scholar] [CrossRef]
- Lu, G.; Zhou, Z.H.; Baker, M.L.; Jakana, J.; Cai, D.; Wei, X.; Chen, S.; Gu, X.; Chiu, W. Structure of double-shelled rice dwarf virus. J. Virol. 1998, 72, 8541–8549. [Google Scholar] [CrossRef]
- Shapiro, A.; Green, T.; Rao, S.; White, S.; Carner, G.; Mertens, P.P.C.; Becnel, J.J. Morphological and molecular characterization of a Cypovirus (Reoviridae) from the mosquito Uranotaenia sapphirina (Diptera: Culicidae). J. Virol. Methods 2005, 79, 9430–9438. [Google Scholar] [CrossRef]
- Brcák, J.; Králík, O. Cytopathic effects of oat sterile dwarf virus in enation cells of oat leaves. Acta Virol. 1980, 24, 346–350. [Google Scholar]
- Day, J.M. The diversity of the orthoreoviruses: Molecular taxonomy and phylogentic divides. Infect. Genet. Evol. 2009, 9, 390–400. [Google Scholar] [CrossRef]
- Bruenn, J.; Madura, K.; Siegel, A.; Miner, Z.; Lee, M. Long internal inverted repeat in a yeast viral double-stranded RNA. Nucleic Acids Res. 1985, 13, 1575–1591. [Google Scholar] [CrossRef]
- Goodman, R.P.; Ghabrial, S.A.; Fichorova, R.N.; Nibert, M.L. Trichomonasvirus: A new genus of protozoan viruses in the family Totiviridae. Arch. Virol. 2011, 156, 171–179. [Google Scholar] [CrossRef]
- Koonin, E.V.; Gorbalenya, A.E. Evolution of RNA genomes: Does the high mutation rate necessitate high rate of evolution of viral proteins? J. Mol. Evol. 1989, 28, 524–527. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V.; Morris, T.J. Evolution and taxonomy of positive-strand RNA viruses: Implications of comparative analysis of amino acid sequences. Crit. Rev. Biochem. Mol. Biol. 1993, 28, 375–430. [Google Scholar] [CrossRef] [PubMed]
- Wolf, Y.I.; Kazlauskas, D.; Iranzo, J.; Lucía-Sanz, A.; Kuhn, J.H.; Krupovic, M.; Dolja, V.V.; Koonin, E.V. Origins and evolution of the global RNA virome. Mbio 2018, 9, e02329-18. [Google Scholar] [CrossRef] [PubMed]
- Mönttinen, H.A.M.; Ravantti, J.J.; Poranen, M.M. Structure unveils relationships between RNA virus polymerases. Viruses 2021, 13, 313. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, S.R.; Wang, D. Origins and challenges of viral dark matter. Virus Res. 2017, 239, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Coatsworth, H.; Bozic, J.; Carrillo, J.; Buckner, E.A.; Rivers, A.R.; Dinglasan, R.R.; Mathias, D.K. Intrinsic variation in the vertically transmitted core virome of the mosquito Aedes aegypti. Mol. Ecol. 2022, 31, 2545–2561. [Google Scholar] [CrossRef]
- Atoni, E.; Zhao, L.; Karungu, S.; Obanda, V.; Agwanda, B.; Xia, H.; Yuan, Z. The discovery and global distribution of novel mosquito-associated viruses in the last decade (2007–2017). Rev. Med. Virol. 2019, 29, e2079. [Google Scholar] [CrossRef]
- Öhlund, P.; Hayer, J.; Lundén, H.; Hesson, J.C.; Blomström, A.L. Viromics reveal a number of novel RNA viruses in Swedish mosquitoes. Viruses 2019, 11, 1027. [Google Scholar] [CrossRef]
- Wang, L.; Rosales Rosas, A.L.; De Coninck, L.; Shi, C.; Bouckaert, J.; Matthijnssens, J.; Delang, L. Establishment of Culex modestus in Belgium and a glance into the virome of Belgian mosquito species. mSphere 2021, 6, e01229-20. [Google Scholar] [CrossRef]
- Peach, D.A.H.; Gries, R.; Young, N.; Lakes, R.; Galloway, E.; Alamsetti, S.K.; Ko, E.; Ly, A.; Gries, G. Attraction of female Aedes aegypti (L.) to aphid honeydew. Insects 2019, 10, 43. [Google Scholar] [CrossRef]
- Peach, D.A.H.; Gries, G. Mosquito phytophagy—Sources exploited, ecological function, and evolutionary transition to haematophagy. Entom. Exp. Appl. 2019, 168, 120–136. [Google Scholar] [CrossRef]
- Beaurepaire, A.L.; Moro, A.; Mondet, F.; Le Conte, Y.; Neumann, P.; Locke, B. Population genetics of ectoparasitic mites suggest arms race with honeybee hosts. Sci. Rep. 2019, 9, 11355. [Google Scholar] [CrossRef]
- Wipf, N.C.; Guidi, V.; Tonolla, M.; Ruinelli, M.; Müller, P.; Engler, O. Evaluation of honey-baited FTA cards in combination with different mosquito traps in an area of low arbovirus prevalence. Parasit Vectors 2019, 12, 554. [Google Scholar] [CrossRef]
- Birnberg, L.; Temmam, S.; Aranda, C.; Correa-Fiz, F.; Talavera, S.; Bigot, T.; Eloit, M.; Busquets, N. Viromics on honey-baited FTA cards as a new tool for the detection of circulating viruses in mosquitoes. Viruses 2020, 12, 274. [Google Scholar] [CrossRef]
- Flies, E.J.; Toi, C.; Weinstein, P.; Doggett, S.L.; Williams, C.R. Converting mosquito surveillance to arbovirus surveillance with honey-baited nucleic acid preservation cards. Vector-Borne Zoonotic Dis. 2015, 15, 397–403. [Google Scholar] [CrossRef]
- Levi, L.I.; Rezelj, V.V.; Henrion-Lacritick, A.; Erazo, D.; Boussier, J.; Vallet, T.; Bernhauerová, V.; Suzuki, Y.; Carrau, L.; Weger-Lucarelli, J.; et al. Defective viral genomes from chikungunya virus are broad-spectrum antivirals and prevent virus dissemination in mosquitoes. PLoS Pathog. 2021, 17, e1009110. [Google Scholar] [CrossRef]
- Rezelj, V.V.; Carrau, L.; Merwaiss, F.; Levi, L.I.; Erazo, D.; Tran, Q.D.; Henrion-Lacritick, A.; Gausson, V.; Suzuki, Y.; Shengjuler, D.; et al. Defective viral genomes as therapeutic interfering particles against flavivirus infection in mammalian and mosquito hosts. Nat. Commun. 2021, 12, 2290. [Google Scholar] [CrossRef]
- Laureti, M.; Paradkar, P.N.; Fazakerley, J.K.; Rodriguez-Andres, J. Superinfection exclusion in mosquitoes and its potential as an arbovirus control strategy. Viruses 2020, 12, 1259. [Google Scholar] [CrossRef]
- He, W.; Chen, Y.; Zhang, X.; Peng, M.; Xu, D.; He, H.; Gao, Y.; Chen, J.; Zhang, J.; Li, Z.; et al. Virome in adult Aedes albopictus captured during different seasons in Guangzhou City, China. Parasites Vectors 2021, 14, 415. [Google Scholar] [CrossRef]
- Feng, Y.; Gou, Q.Y.; Yang, W.H.; Wu, W.C.; Wang, J.; Holmes, E.C.; Liang, G.; Shi, M. A time-series meta-transcriptomic analysis reveals the seasonal, host, and gender structure of mosquito viromes. Virus Evol. 2022, 8, veac006. [Google Scholar] [CrossRef]
- Shi, C.; Zhao, L.; Atoni, E.; Zeng, W.; Hu, X.; Matthijnssens, J.; Yuan, Z.; Xia, H. Stability of the virome in lab- and field-collected Aedes albopictus mosquitoes across different developmental stages and possible core viruses in the publicly available virome data of Aedes mosquitoes. mSystems 2020, 5, e00640-20. [Google Scholar] [CrossRef]
- Kubacki, J.; Flacio, E.; Qi, W.; Guidi, V.; Tonolla, M.; Fraefel, C. Viral metagenomic analysis of Aedes albopictus mosquitos from southern Switzerland. Viruses 2020, 12, 929. [Google Scholar] [CrossRef]
- Traavik, T.; Mehl, R.; Wiger, R. California encephalitis group viruses isolated from mosquitoes collected in Southern and Arctic Norway. Acta Path. Microbiol. Scand. Sect. B 1978, 86B, 335–342. [Google Scholar] [CrossRef]
- Putkuri, N.; Vaheri, A.; Vapalahti, O. Prevalence and protein specificity of human antibodies to Inkoo virus infection. Clin. Vaccine Immunol. 2007, 14, 1555–1562. [Google Scholar] [CrossRef]
- Xia, H.; Wang, Y.; Shi, C.; Atoni, E.; Zhao, L.; Yuan, Z. Comparative metagenomic profiling of viromes associated with four common mosquito species in China. Virol. Sin. 2018, 33, 59–66. [Google Scholar] [CrossRef]
- Hameed, M.; Wahaab, A.; Shan, T.; Wang, X.; Khan, S.; Di, D.; Xiqian, L.; Zhang, J.J.; Anwar, M.N.; Nawaz, M.; et al. A metagenomic analysis of mosquito virome collected from different animal farms at Yunnan-Myanmar border of China. Front. Microbiol. 2020, 11, 591478. [Google Scholar] [CrossRef]
- Oguzie, J.U.; Nwangwu, U.C.; Oluniyi, P.E.; Olumade, T.J.; George, U.E.; Kazeem, A.; Bankole, B.E.; Brimmo, F.O.; Asadu, C.C.; Chukwuekezie, O.C.; et al. Metagenomic sequencing characterizes a wide diversity of viruses in field mosquito samples in Nigeria. Sci. Rep. 2022, 12, 7616. [Google Scholar] [CrossRef]
- Shi, C.; Beller, L.; Deboutte, W.; Yinda, K.C.; Delang, L.; Vega-Rúa, A.; Failloux, A.-B.; Matthijnssens, J. Stable distinct core eukaryotic viromes in different mosquito species from Guadeloupe, using single mosquito viral metagenomics. Microbiome 2019, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Hameed, M.; Liu, K.; Anwar, M.N.; Wahaab, A.; Li, C.; Di, D.; Wang, X.; Khan, S.; Xu, J.; Li, B.; et al. A viral metagenomic analysis reveals rich viral abundance and diversity in mosquitoes from pig farms. Transbound Emerg. Dis. 2020, 67, 328–343. [Google Scholar] [CrossRef] [PubMed]
- Nebbak, A.; Monteil-Bouchard, S.; Berenger, J.-M.; Almeras, L.; Parola, P.; Desnues, C. Virome diversity among mosquito populations in a sub-urban region of Marseille, France. Viruses 2021, 13, 768. [Google Scholar] [CrossRef] [PubMed]
- Viljakainen, L.; Borshagovski, A.-M.; Saarenpää, S.; Kaitala, A.; Jurvansuu, J. Identification and characterisation of common glow-worm RNA viruses. Virus Genes 2020, 56, 236–248. [Google Scholar] [CrossRef]
- Kurkela, S.; Manni, T.; Myllynen, J.; Vaheri, A.; Vapalahti, O. Clinical and laboratory manifestations of Sindbis virus infection: Prospective study, Finland, 2002–2003. J. Infect. Dis. 2005, 191, 1820–1829. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truong Nguyen, P.T.; Culverwell, C.L.; Suvanto, M.T.; Korhonen, E.M.; Uusitalo, R.; Vapalahti, O.; Smura, T.; Huhtamo, E. Characterisation of the RNA Virome of Nine Ochlerotatus Species in Finland. Viruses 2022, 14, 1489. https://doi.org/10.3390/v14071489
Truong Nguyen PT, Culverwell CL, Suvanto MT, Korhonen EM, Uusitalo R, Vapalahti O, Smura T, Huhtamo E. Characterisation of the RNA Virome of Nine Ochlerotatus Species in Finland. Viruses. 2022; 14(7):1489. https://doi.org/10.3390/v14071489
Chicago/Turabian StyleTruong Nguyen, Phuoc T., C. Lorna Culverwell, Maija T. Suvanto, Essi M. Korhonen, Ruut Uusitalo, Olli Vapalahti, Teemu Smura, and Eili Huhtamo. 2022. "Characterisation of the RNA Virome of Nine Ochlerotatus Species in Finland" Viruses 14, no. 7: 1489. https://doi.org/10.3390/v14071489