Small Body, Large Chromosomes: Centric Fusions Shaped the Karyotype of the Amazonian Miniature Fish Nannostomus anduzei (Characiformes, Lebiasinidae)

 ,
,  ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Chromosomal Preparation
2.2. Conventional and Molecular Cytogenetics
2.3. Microscopic Analyses and Image Processing
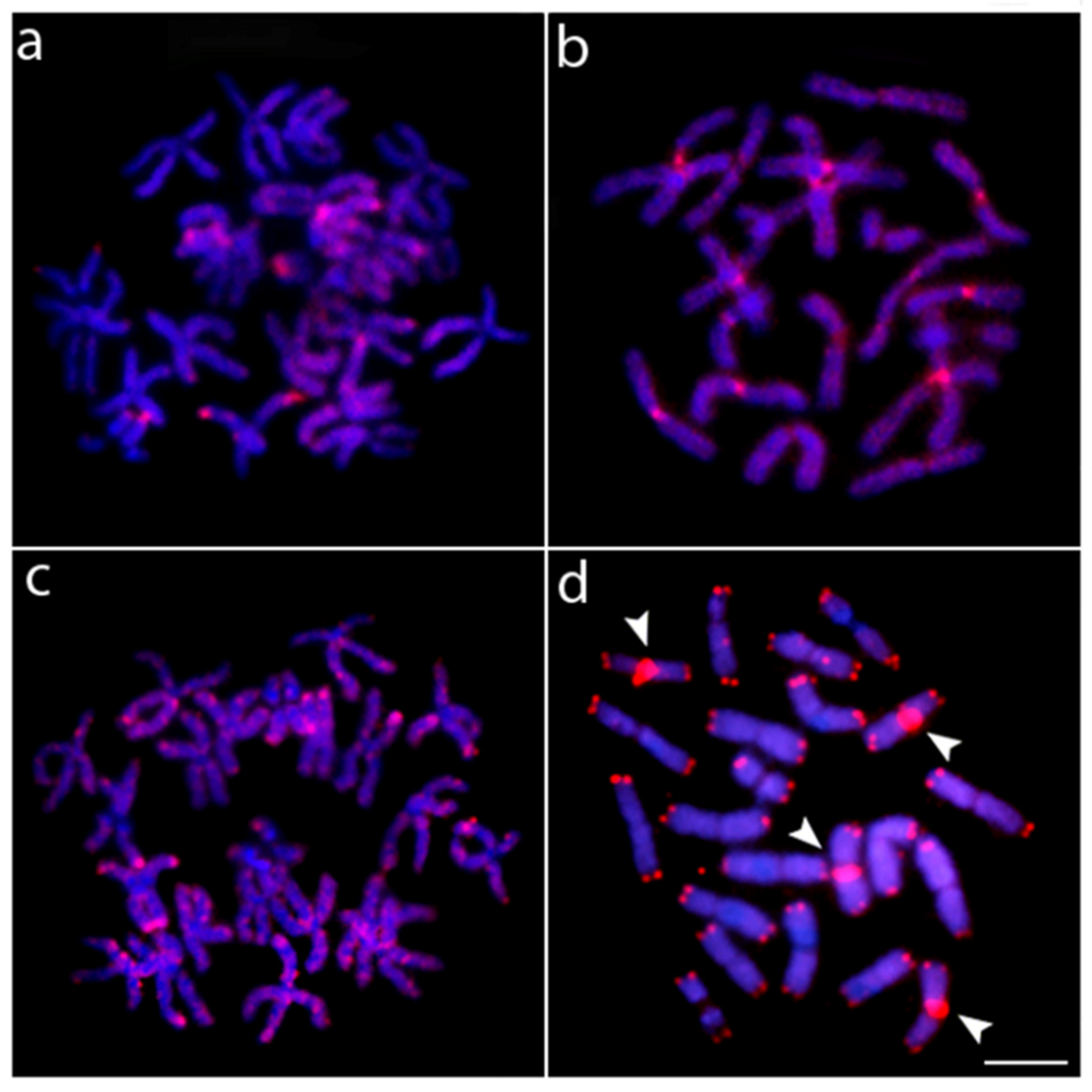
3. Results
4. Discussion
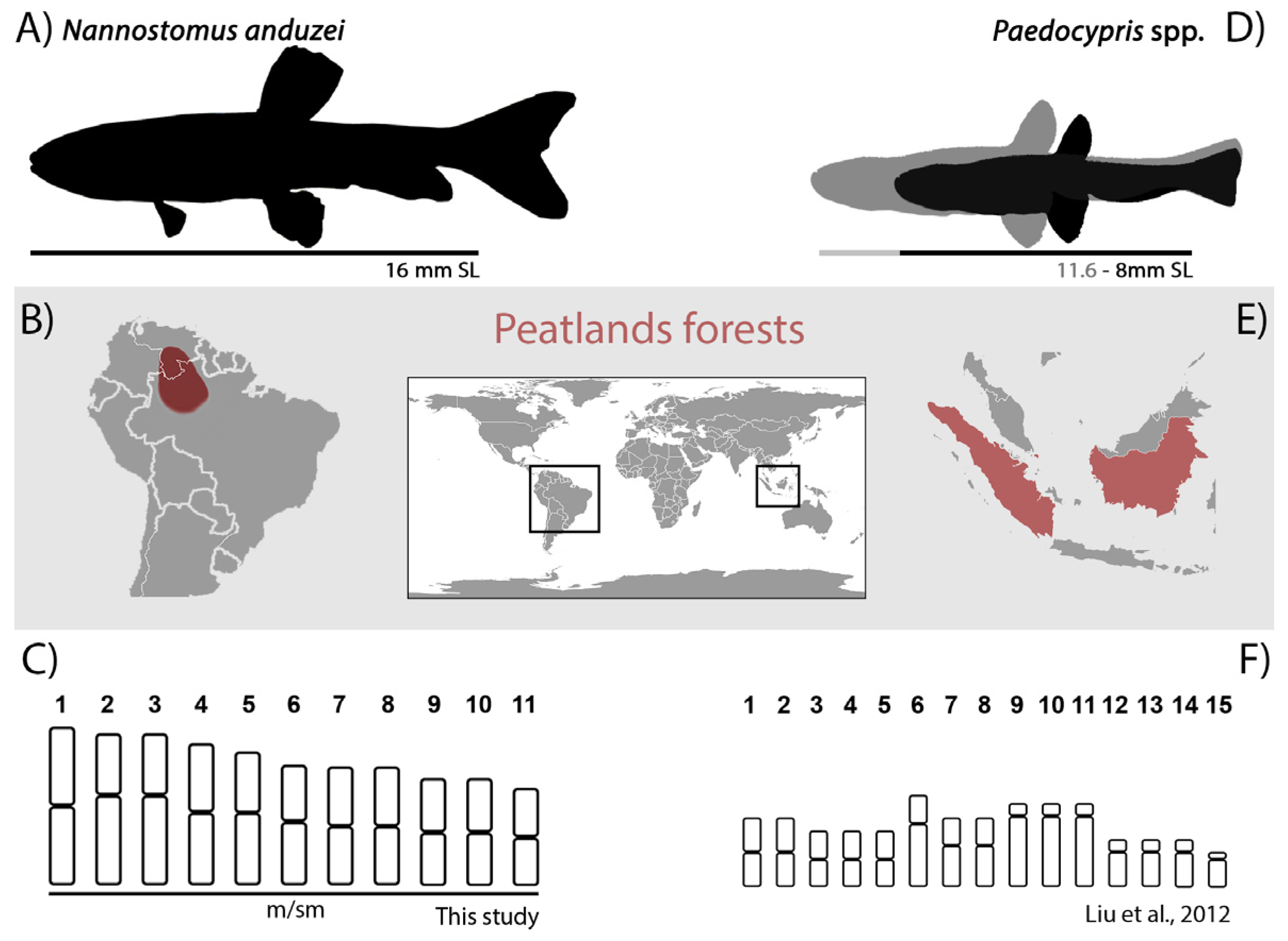
4.1. Chromosomal Characteristics of Nannostomus Anduzei
4.2. Chromosomal Reduction and Miniaturization: Cause-Effect Relationship or an Indirect Association?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanken, J.; Wake, D.B. Miniaturization of body size: Organismal consequences and evolutionary significance. Annu. Rev. Ecol. Syst. 1993, 24, 501–519. [Google Scholar] [CrossRef]
- Rundell, R.J.; Leander, B.S. Masters of miniaturization: Convergent evolution among interstitial eukaryotes. Bioessays 2010, 32, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, S.H.; Vari, R.P. Miniaturization in South American freshwater fishes: An overview and discussion. Proc. Biol. Soc. Washingt. 1988, 101, 444–465. [Google Scholar]
- Kottelat, M.; Vidthayanon, C. Boraras micros, a new genus and species of minute freshwater fish from Thailand (Teleostei: CYprinidae). Ichthyol. Explor. Freshw. 1993, 4, 161–176. [Google Scholar]
- Conway, K.W.; Moritz, T. Barboides britzi, a new species of miniature cyprinid from Benin (Ostariophysi: Cyprinidae), with a neotype designation for B. gracilis. Ichthyol. Explor. Freshw. 2006, 17, 73–84. [Google Scholar]
- Bennett, M.G.; Conway, K.W. An overview of North America's diminutive freshwater fish fauna. Ichthyol. Explor. Freshw. 2010, 21, 63–72. [Google Scholar]
- Marinho, M.M.F.; Bastos, D.A.; Menezes, N.A. New species of miniature fish from Marajó Island, Pará, Brazil, with comments on its relationships (Characiformes: Characidae). Neotrop. Ichthyol. 2013, 11, 739–746. [Google Scholar] [CrossRef] [Green Version]
- Toledo-Piza, M.; Mattox, G.M.T.; Britz, R. Priocharax nanus, a new miniature characid from the rio Negro, Amazon basin (Ostariophysi: Characiformes), with an updated list of miniature Neotropical freshwater fishes. Neotrop. Ichthyol. 2014, 12, 229–246. [Google Scholar] [CrossRef] [Green Version]
- Abrahão, V.P.; Pastana, M.; Marinho, M. On a remarkable sexual dimorphic trait in the Characiformes related to the olfactory organ and description of a new miniature species of Tyttobrycon Géry (Characiformes: Characidae). PLoS ONE 2019, 14, e0226130. [Google Scholar] [CrossRef] [Green Version]
- Mattox, G.M.T.; Souza, C.S.; Toledo-Piza, M.; Britz, R.; Oliveira, C. A new miniature species of Priocharax (Teleostei: Characiformes: Characidae) from the rio Madeira drainage, Brazil, with comments on the adipose fin in Characiforms. Verteb Zool. 2020, 70, 417–433. [Google Scholar] [CrossRef]
- Mattox, G.M.T.; Souza, C.S.; Toledo-Piza, M.; Oliveira, C. A new miniature species of Priocharax (Characiformes: Characidae) from the upper rio Ipixuna, Purus drainage, Brazil. Neotrop. Ichthyol. 2021, 19, e210048. [Google Scholar] [CrossRef]
- Rodrigues, E.K.D.Q.; Netto-Ferreira, A.L. A new miniature species of Odontocharacidium (Characiformes: Crenuchidae) from the Río Orinoco basin, Venezuela. Neotrop. Ichthyol. 2020, 18, e190008. [Google Scholar] [CrossRef]
- Perkin, J.S.; Montaña, C.G.; Nogueira, E.J.; Brandão, B.B.; Mattox, G.M.; Conway, K.W. Estimated richness and environmental correlates of miniature fish assemblages in the rio Jacundá, Brazil. Neotrop. Ichthyol. 2022, 20, e210051. [Google Scholar] [CrossRef]
- Ravi, V.; Venkatesh, B. Rapidly evolving fish genomes and teleost diversity. Curr. Opin. Genet. Dev. 2008, 18, 544–550. [Google Scholar] [CrossRef]
- Hufton, A.L.; Groth, D.; Vingron, M.; Lehrach, H.; Poustka, A.J.; Panopoulou, G. Early vertebrate whole genome duplications were predated by a period of intense genome rearrangement. Genome Res. 2008, 18, 1582–1591. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Ye, X.; Yang, Y.; Yang, Y.; Sun, Y.H.; Mei, Y.; Xiong, S.; He, K.; Xu, L.; Fang, Q. Comparative genomics sheds light on the convergent evolution of miniaturized wasps. Mol. Biol. Evolution. 2021, 38, 5539–5554. [Google Scholar] [CrossRef] [PubMed]
- Sessions, S.K.; Larson, A. Developmental correlates of genome size in plethodontid salamanders and their implications for genome evolution. Evolution 1987, 41, 1239–1251. [Google Scholar] [CrossRef]
- Roth, G.; Rottluff, B.; Grunwald, W.; Hanken, J.; Linke, R. Miniaturization in plethodontid salamanders (Caudata: Plethodontidae) and its consequences for the brain and visual system. Biol. J. Linn. Soc. 1990, 40, 165–190. [Google Scholar] [CrossRef]
- Liu, S.; Hui, T.H.; Tan, S.L.; Hong, Y. Chromosome evolution and genome miniaturization in minifish. PLoS ONE 2012, 7, e37305. [Google Scholar] [CrossRef] [Green Version]
- Malmstrøm, M.; Britz, R.; Matschiner, M.; Tørresen, O.K.; Hadiaty, R.K.; Yaakob, N.; Tan, H.H.; Jakobsen, K.S.; Salzburger, W.; Rüber, L. The most developmentally truncated fishes show extensive Hox gene loss and miniaturized genomes. Genome Biol. Evol. 2018, 10, 1088–1103. [Google Scholar] [CrossRef]
- Weitzman, M.; Weitzman, S.H. Family Lebiasinidae. In Check List of the Freshwater Fishes of South and Central America; Edipucrs: Porto Alegre, Brazil, 2003; pp. 241–250. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 25 October 2022).
- Sember, A.; de Oliveira, E.A.; Ráb, P.; Bertollo, L.A.C.; de Freitas, N.L.; Viana, P.F.; Yano, C.F.; Hatanaka, T.; Marinho, M.M.F.; Moraes, R.L.R.; et al. Centric Fusions behind the Karyotype Evolution of Neotropical Nannostomus Pencilfishes (Characiforme, Lebiasinidae): First Insights from a Molecular Cytogenetic Perspective. Genes 2020, 11, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheel, J.J. Fish Chromosomes and Their Evolution; Danmarks Akvarium: Charlottenlund, Denmark, 1973. [Google Scholar]
- Oliveira, C.; Andreata, A.A.; Almeida-Toledo, L.F.; Toledo-Filho, S.A. Karyotype and nucleolus organizer regions of Pyrrhulina cf. australis (Pisces, Characiformes, Lebiasinidae). Rev. Bras. Genet. 1991, 14, 685–690. [Google Scholar]
- Arai, R. Fish Karyotypes: A Check List; Springer Science & Business Media: Tokyo, Japan, 2011. [Google Scholar]
- Moraes, R.L.R.; Bertollo, L.A.C.; Marinho, M.M.F.; Yano, C.F.; Hatanaka, T.; Barby, F.F.; Troy, W.P.; Cioffi, M.B. Evolutionary relationships and cytotaxonomy considerations in the genus Pyrrhulina (Characiformes, Lebiasinidae). Zebrafish 2017, 14, 536–546. [Google Scholar] [CrossRef]
- Moraes, R.L.R.; Sember, A.; Bertollo, L.A.C.; de Oliveira, E.A.; Ráb, P.; Hatanaka, T.; Marinho, M.M.F.; Liehr, T.; Al-Rikabi, A.B.H.; Feldberg, E.; et al. Comparative cytogenetics and neo-Y formation in small-sized fish species of the genus Pyrrhulina (Characiformes, Lebiasinidae). Front. Genet. 2019, 10, 678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraes, R.L.R.; Sassi, F.D.M.C.; Bertollo, L.A.C.; Marinho, M.M.F.; Viana, P.F.; Feldberg, E.; Oliveira, V.C.S.; Deon, G.A.; Al-Rikabi, A.B.H.; Liehr, T.; et al. Tracking the evolutionary trends among small-size fishes of the genus Pyrrhulina (Characiforme, Lebiasinidae): New insights from a molecular cytogenetic perspective. Front. Genet. 2021, 12, 769984. [Google Scholar] [CrossRef]
- Sassi, F.M.C.; de Oliveira, E.A.; Bertollo, L.A.C.; Nirchio, M.; Hatanaka, T.; Marinho, M.M.F.; Moreira-Filho, O.; Aroutiounian, R.; Liehr, T.; Al-Rikabi, A.B.H.; et al. Chromosomal evolution and evolutionary relationships of Lebiasina species (Characiformes, Lebiasinidae). Int. J. Mol. Sci. 2019, 20, 2944. [Google Scholar] [CrossRef] [Green Version]
- Sassi, F.M.C.; Hatanaka, T.; Moraes, R.L.R.; Toma, G.A.; Oliveira, E.A.; Liehr, T.; Ráb, P.; Bertollo, L.A.C.; Viana, P.E.; Felberg, E.; et al. An Insight into the Chromosomal Evolution of Lebiasinidae (Teleostei, Characiformes). Genes 2020, 11, 365. [Google Scholar] [CrossRef]
- Toma, G.A.; Moraes, R.L.R.; Sassi, F.M.C.; Bertollo, L.A.C.; de Oliveira, E.A.; Ráb, P.; Sember, A.; Liehr, T.; Hatanaka, T.; Viana, P.F.; et al. Cytogenetics of the small-sized fish, Copeina guttata (Characiformes, Lebiasinidae): Novel insights into the karyotype differentiation of the family. PLoS ONE 2019, 14, e0226746. [Google Scholar] [CrossRef]
- Bertollo, L.A.C.; Cioffi, M.B.; Moreira-Filho, O. Direct chromosome preparation from Freshwater teleost fishes. In Fish Cytogenetic Techniques (Ray-Fin Fishes and Chondrichthyans), 1st ed.; Ozouf-Costaz, C., Pisano, E., Foresti, F., Toledo, L.F.A., Eds.; CRC Press: Boca Raton, FL, USA, 2015; Volume 1, pp. 21–26. [Google Scholar]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Yano, C.F.; Bertollo, L.A.C.; Cioffi, M.B. Fish-FISH: Molecular cytogenetics in fish species. In Fluorescence in Situ Hybridization (FISH)—Application Guide, 2nd ed.; Liehr, T., Ed.; Springer: Berlin, Germany, 2017; pp. 429–444. [Google Scholar]
- Pendás, A.M.; Moran, P.; Freije, J.P.; Garcia-Vazquez, E. Chromosomal Mapping and Nucleotide Sequence of Two Tandem Repeats of Atlantic salmon 5S rDNA. Cytogenet. Cel Genet. 1994, 67, 31–36. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Martins, C.; Centofante, L.; Jacobina, U.; Bertollo, L.A.C. Chromosomal variability among allopatric populations of Erythrinidae fish Hoplias malabaricus: Mapping of three classes of repetitive DNAs. Cytogenet. Genome Res. 2009, 125, 132–141. [Google Scholar] [CrossRef]
- Kubat, Z.; Hobza, R.; Vyskot, B.; Kejnovsky, E. Microsatellite Accumulation on the Y Chromosome in Silene latifolia. Genome 2008, 51, 350–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved Telomere Detection Using a Telomere Repeat Probe (TTAGGG)n generated by PCR. Nucl. Acids Res. 1991, 19, 4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Marinho, M.M.F. Relações filogenéticas e revisão taxonômica das espécies do gênero Copella Myers, 1956 (Characiformes: Lebiasinidae). Ph.D. Thesis, Universidade Estadual Paulista Julio de Mesquita Filho, São Paulo, Brazil, 2014. [Google Scholar]
- Netto-Ferreira, A.L. Revisão taxonômica e relações interespecíficas de Lebiasininae (Ostariophysi: Characiformes: Lebiasinidae). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2010. [Google Scholar]
- Bolzán, A.D. Interstitial telomeric sequences in vertebrate chromosomes: Origin, function, instability and evolution. Mutat. Res. /Rev. Mutat. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef]
- Barra, V.; Fachinetti, D. The dark side of centromeres: Type, causes, and consequences of structural abnormalities implicating centromeric DNA. Nat Commun. 2018, 9, 4340. [Google Scholar] [CrossRef] [Green Version]
- Garagna, S.D.; Broccoli, C.A.; Redi, J.B.; Searle, H.J.; Cooke, E. Robertisonian metacentrics of the house mouse lose telomeric sequences but retain some minor satellite DNA in the pericentromeric area. Chromosoma 1995, 103, 685–692. [Google Scholar] [CrossRef]
- Nanda, I.S.; Schneider-Rasp, H.; Winking, M. Schmid Loss of telomeric sites in the chromosomes of Mus musculus domesticus (Rodentia: Muridae) during Robertsonian rearrangements. Chromosome Res. 1995, 3, 399–409. [Google Scholar] [CrossRef]
- Glugoski, L.; Deon, G.; Schott, S.; Vicari, M.R.; Nogaroto, V.; Moreira-Filho, O. Comparative cytogenetic analyses in Ancistrus species (Siluriformes: Loricariidae). Neotrop. Ichthyol. 2020, 18, e200013. [Google Scholar] [CrossRef]
- Guzmán-Markevich, K.; Roco, Á.S.; Ruiz-García, A.; Bullejos, M. Cytogenetic Analysis in the Toad Species Bufo spinosus, Bufotes viridis and Epidalea calamita (Anura, Bufonidae) from the Mediterranean Area. Genes 2022, 13, 1475. [Google Scholar] [CrossRef]
- Cappelletti, E.; Piras, F.M.; Sola, L.; Santagostino, M.; Abdelgadir, W.A.; Raimondi, E.; Lescai, F.; Nergadze, S.G.; Giulotto, E. Robertsonian fusion and centromere repositioning contributed to the formation of satellite-free centromeres during the evolution of zebras. Mol. Biol. Evol. 2022, 39, msac162. [Google Scholar] [CrossRef]
- Ocalewicz, K. Telomeres in fishes. Cytogenet. Genome Res. 2013, 141, 114–125. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Martins, C.; Bertollo, L.A.C. Chromosome spreading of associated transposable elements and ribosomal DNA in the fish Erythrinus erythrinus. Implications for genome change and karyoevolution in fish. BMC Evol Biol. 2010, 10, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cioffi, M.B.; Bertollo, L.A.C. Initial steps in XY chromosome differentiation in Hoplias malabaricus and the origin of an X(1)X(2)Y sex chromosome system in this fish group. Heredity 2010, 105, 554–561. [Google Scholar] [CrossRef] [Green Version]
- Sola, L.; Gornung, E.; Naoi, H.; Gunji, R.; Sato, C.; Kawamura, K.; Arai, R.; Ueda, T. FISH-mapping of 18S ribosomal RNA genes and telomeric sequences in the Japanese bitterlings Rhodeus ocellatus kurumeus and Tanakia limbata (Pisces, Cyprinidae) reveals significant cytogenetic differences in morphologically similar karyotypes. Genetica 2003, 119, 99–106. [Google Scholar] [CrossRef]
- Blanco, D.R.; Vicari, M.R.; Lui, R.L.; Bertollo, L.A.C.; Traldi, J.B.; Moreira-Filho, O. The role of the Robertsonian rearrangements in the origin of the XX/XY 1 Y 2 sex chromosome system and in the chromosomal differentiation in Harttia species (Siluriformes, Loricariidae). Rev. Fish Biol. Fish. 2012, 23, 127–134. [Google Scholar] [CrossRef]
- De Marco Ferro, D.A.; Moreira-Filho, O.; Bertollo, L.A.C. B chromosome polymorphism in the fish, Astyanax scabripinnis. Genetica 2003, 119, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Gromicho, M.; Coutanceau, J.P.; Ozouf-Costaz, C.; Collares-Pereira, M.J. Contrast between extensive variation of 28S rDNA and stability of 5S rDNA and telomeric repeats in the diploid-polyploid Squalius alburnoides complex and in its maternal ancestor Squalius pyrenaicus (Teleostei, Cyprinidae). Chromosome Res. 2006, 14, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Su, Y.; Ding, S.; Cai, Y.; Wang, J. Cytogenetic analysis of Orange spotted grouper, Epinephelus coioides, using chromosome banding and fluorescence in situ hybridization. Hydrobiologia 2010, 638, 1–10. [Google Scholar] [CrossRef]
- Malimpensa, G.C.; Traldi, J.B.; Toyama, D.; Henrique-Silva, F.; Vicari, M.R.; Moreira-Filho, O. Chromosomal mapping of repeat DNA in Bergiaria westermanni (Pimelodidae, Siluriformes): Localization of 45S rDNA in B chromosomes. Cytogenet. Genome Res. 2018, 154, 99–106. [Google Scholar] [CrossRef]
- Soares, L.B.; Paim, F.G.; Ramos, L.P.; Foresti, F.; Oliveira, C. Molecular cytogenetic analysis and the establishment of a cell culture in the fish species Hollandichthys multifasciatus (Eigenmann & Norris, 1900) (Characiformes, Characidae). Genet. Mol. Biol. 2021, 44, e20200260. [Google Scholar] [CrossRef] [PubMed]
- Barros, A.V.; Wolski, M.A.V.; Nogaroto, V.; Almeida, M.C.; Moreira-Filho, O.; Vicari, M.R. Fragile sites, dysfunctional telomere and chromosome fusions: What is 5S rDNA role? Gene 2017, 608, 20–27. [Google Scholar] [CrossRef]
- Cavalcante, M.G.; Eduardo, C.; Carvalho, M.; Nagamachi, Y.; Pieczarka, J.C.; Vicari, M.R.; Noronha, R.C.R. Physical mapping of repetitive DNA suggests 2n reduction in Amazon turtles Podocnemis (Testudines: Podocnemididae). PLoS ONE 2018, 13, e0197536. [Google Scholar] [CrossRef] [Green Version]
- Glugoski, L.; Giuliano-Caetano, L.; Moreira-Filho, O.; Vicari, M.R.; Nogaroto, V. Co-located hAT transposable element and 5S rDNA in an interstitial telomeric sequence suggest the formation of Robertsonian fusion in armored catfish. Gene 2018, 650, 49–54. [Google Scholar] [CrossRef]
- Leite, P.P.D.M.; Sassi, F.D.M.C.; Marinho, M.M.F.; Nirchio, M.; Moraes, R.L.R.; Toma, G.A.; Bertollo, L.A.C.; Cioffi, M.B. Tracking the evolutionary pathways among Brazilian Lebiasina species (Teleostei: Lebiasinidae): A chromosomal and genomic comparative investigation. Neotrop. Ichthyol. 2022, 20, e210153. [Google Scholar] [CrossRef]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Symonová, R.; Majtánová, Z.; Sember, A.; Staaks, G.B.; Bohlen, J.; Freyhof, J.; Rábová, M.; Ráb, P. Genome differentiation in a species pair of coregonine fishes: An extremely rapid speciation driven by stress-activated retrotransposons mediating extensive ribosomal DNA multiplications. BMC Evol. Biol. 2013, 13, 42. [Google Scholar] [CrossRef] [Green Version]
- Sember, A.; Bohlen, J.; Šlechtová, V.; Altmanová, M.; Symonová, R.; Ráb, P. Karyotype differentiation in 19 species of river loach fishes (Nemacheilidae, Teleostei): Extensive variability associated with rDNA and heterochromatin distribution and its phylogenetic and ecological interpretation. BMC Evol. Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [Green Version]
- Symonová, R.; Howell, W.M. Vertebrate genome evolution in the light of fish cytogenomics and rDNAomics. Genes 2018, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Maneechot, N.; Yano, C.F.; Bertollo, L.A.C.; Getlekha, N.; Molina, W.F.; Ditcharoen, S.; Tengjaroenkul, B.; Supiwong, W.; Tanomtong, A.; Cioffi, M.B. Genomic organization of repetitive DNAs highlights chromosomal evolution in the genus Clarias (Clariidae, Siluriformes). Molec. Cytogenet. 2016, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- da Silva, F.A.; Feldberg, E.; Carvalho, N.D.M.; Rangel, S.M.H.R.; Schneider, C.H.; Carvalho-Zilse, G.A.; da Silva, V.F.; Gross, M.C. Effects of environmental pollution on the rDNAomics of Amazonian fish. Environ. Pollut. 2019, 252, 180–187. [Google Scholar] [CrossRef]
- Jernfors, T.; Danforth, J.; Kesäniemi, J.; Lavrinienko, A.; Tukalenko, E.; Fajkus, J.; Dvořáčková, M.; Mappes, T.; Watts, P.C. Expansion of rDNA and pericentromere satellite repeats in the genomes of bank voles Myodes glareolus exposed to environmental radionuclides. Ecol. Evol. 2021, 11, 8754–8767. [Google Scholar] [CrossRef]
- Kobayashi, T. Ribosomal RNA gene repeats, their stability and cellular senescence. Proc. Jpn. Acad. 2014, 90, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calcagnotto, D.; Schaefer, S.A.; DeSalle, R. Relationships among characiform fishes inferred from analysis of nuclear and mitochondrial gene sequences. Mol. Phylogenet. Evol. 2005, 36, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Bertollo, L.A.C. Chromosomal distribution and evolution of repetitive DNAs in fish. In Repetitive DNA; Garrido-Ramos, M.A., Ed.; Karger: Basel, Switzerland, 2012; pp. 197–221. [Google Scholar] [CrossRef]
- Pokorná, M.; Kratochvíl, L.; Kejnovský, E. Microsatellite distribution on sex chromosomes at different stages of heteromorphism and heterochromatinization in two lizard species (Squamata: Eublepharidae: Coleonyx elegans and Lacertidae: Eremias velox). BMC Genetics. 2011, 12, 90. [Google Scholar] [CrossRef] [Green Version]
- Kejnovský, E.; Michalovova, M.; Steflova, P.; Kejnovska, I.; Manzano, S.; Hobza, R.; Kubat, Z.; Kovarik, J.; Jamilena, M.; Vyskot, B. Expansion of microsatellites on evolutionary young Y chromosome. PLoS ONE 2013, 8, e45519. [Google Scholar] [CrossRef] [Green Version]
- Poltronieri, J.; Marquioni, V.; Bertollo, L.A.C.; Kejnovsky, E.; Molina, W.F.; Liehr, T.; Cioffi, M.B. Comparative Chromosomal Mapping of Microsatellites in Leporinus Species (Characiformes, Anostomidae): Unequal Accumulation on the W Chromosomes. Cytogenet. Genome Res. 2014, 142, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.C.; de Oliveira, E.A.; Bertollo, L.A.C.; Yano, C.F.; Oliveira, C.; Decru, E.; Jegede, O.I.; Hatanaka, T.; Liehr, T.; Al-Rikabi, A.B.H.; et al. First chromosomal analysis in Hepsetidae (Actinopterygii, Characiformes): Insights into relationship between African and Neotropical fish groups. Front. Genet. 2017, 8, 203. [Google Scholar] [CrossRef] [Green Version]
- Ditcharoen, S.; Sassi, F.M.C.; Bertollo, L.A.C.; Molina, W.F.; Liehr, T.; Saenjundaeng, P.; Tanomtong, A.; Supiwong, W.; Suwannapoom, C.; Cioffi, M.B. Comparative Chromosomal Mapping of Microsatellite Repeats Reveals Divergent Patterns of Accumulation in 12 Siluridae (Teleostei: Siluriformes) Species. Genet. Mol. Biol. 2020, 43, e20200091. [Google Scholar] [CrossRef] [PubMed]
- Kottelat, M.; Britz, R.; Hui, T.H.; Witte, K.E. Paedocypris, a new genus of Southeast Asian cyprinid fish with a remarkable sexual dimorphism, comprises the world's smallest vertebrate. Proc. R. Soc. B Biol. Sci. 2006, 273, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Rüber, L.; Kottelat, M.; Tan, H.H.; Ng, P.K.; Britz, R. Evolution of miniaturization and the phylogenetic position of Paedocypris, comprising the world's smallest vertebrate. BMC Evol. Biol. 2007, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Gumbricht, T.; Roman-Cuesta, R.M.; Verchot, L.; Herold, M.; Wittmann, F.; Householder, E.; Herold, N.; Murdiyarso, D. An expert system model for mapping tropical wetlands and peatlands reveals South America as the largest contributor. Global Chang. Biology 2017, 23, 3581–3599. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, K.; Sado, T.; Mayden, R.L.; Hanzawa, N.; Nakamura, K.; Nishida, M.; Miya, M. Mitogenomic Evolution and Interrelationships of the Cypriniformes (Actinopterygii: Ostariophysi): The First Evidence Toward Resolution of Higher-Level Relationships of the World’s Largest Freshwater Fish Clade Based on 59 Whole Mitogenome Sequences. J. Mol. Evol. 2006, 63, 826–841. [Google Scholar] [CrossRef]
- Mayden, R.L.; Chen, W.J. The world’s smallest vertebrate species of the genus Paedocypris: A new family of freshwater fishes and the sister group to the world’s most diverse clade of freshwater fishes (Teleostei: Cypriniformes). Mol. Phylogenetics Evol. 2010, 57, 152–175. [Google Scholar] [CrossRef]
- Sam, K.K.; Lau, N.S.; Shu-Chien, C.A.; Muchlisin, Z.A.; Nugrobo, R.A. Complete Mitochondrial Genomes of Paedocypris micromegethes and Paedocypris carbunculus Reval Conserved gene Order and Phylogenetic relationships of Miniaturized Cyprinids. Front. Ecol. Evol. 2021, 9, 662501. [Google Scholar] [CrossRef]
- Supiwong, W.; Pinthong, K.; Seetapan, K.; Saenjundaeng, P.; Bertollo, L.A.C.; de Oliveira, E.A.; Yano, C.F.; Liehr, T.; Phimphan, S.; Tamomtong, A.; et al. Karyotype diversity and evolutionary trends in the Asian swamp eel Monopterus albus (Synbranchiformes, Synbranchidae): A case of chromosomal speciation? BMC Evol Biol. 2019, 19, 73. [Google Scholar] [CrossRef] [Green Version]
- Amores, A.; Wilson, C.A.; Allard, C.A.H.; Detrich, H.W., III; Postlethwait, J.H. Cold Fusion: Massive Karyotype Evolution in the Antarctic Bullhead Notothen Notothenia coriiceps. G3 Genes Genomes Genet. 2017, 7, 2195–2207. [Google Scholar] [CrossRef] [Green Version]
- Walsh, J.B. Rate of accumulation of reproductive isolation by chromosome rearrangements. Am. Nat. 1982, 120, 510–532. [Google Scholar] [CrossRef]
- Phillips, R.; Rab, P. Chromosome evolution in the Salmonidae (Pisces): An update. Biol. Rev. 2001, 76, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Qumsiyeh, M.B. Evolution of number and morphology of mammalian chromosomes. J. Hered. 1994, 85, 455–465. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Moraes, R.L.R.; Sassi, F.d.M.C.; Marinho, M.M.F.; Ráb, P.; Porto, J.I.R.; Feldberg, E.; Cioffi, M.d.B. Small Body, Large Chromosomes: Centric Fusions Shaped the Karyotype of the Amazonian Miniature Fish Nannostomus anduzei (Characiformes, Lebiasinidae). Genes 2023, 14, 192. https://doi.org/10.3390/genes14010192
de Moraes RLR, Sassi FdMC, Marinho MMF, Ráb P, Porto JIR, Feldberg E, Cioffi MdB. Small Body, Large Chromosomes: Centric Fusions Shaped the Karyotype of the Amazonian Miniature Fish Nannostomus anduzei (Characiformes, Lebiasinidae). Genes. 2023; 14(1):192. https://doi.org/10.3390/genes14010192
Chicago/Turabian Stylede Moraes, Renata Luiza Rosa, Francisco de Menezes Cavalcante Sassi, Manoela Maria Ferreira Marinho, Petr Ráb, Jorge Ivan Rebelo Porto, Eliana Feldberg, and Marcelo de Bello Cioffi. 2023. "Small Body, Large Chromosomes: Centric Fusions Shaped the Karyotype of the Amazonian Miniature Fish Nannostomus anduzei (Characiformes, Lebiasinidae)" Genes 14, no. 1: 192. https://doi.org/10.3390/genes14010192