
Changes in the Immunity, Histopathology, and Metabolism of Crayfish (Procambarus clarkii) in Response to Drought


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Drought Exposure
2.2. Total Hemocytes Count
2.3. Phagocytosis Assay
2.4. ROS Detection
2.5. Hematoxylin and Eosin Staining
2.6. Metabolomics Analysis
2.6.1. Metabolite Extraction
2.6.2. Liquid Chromatograph Conditions
2.6.3. Mass Spectrometry Conditions
2.6.4. Statistical Analysis
2.7. Statistical Analysis
3. Results
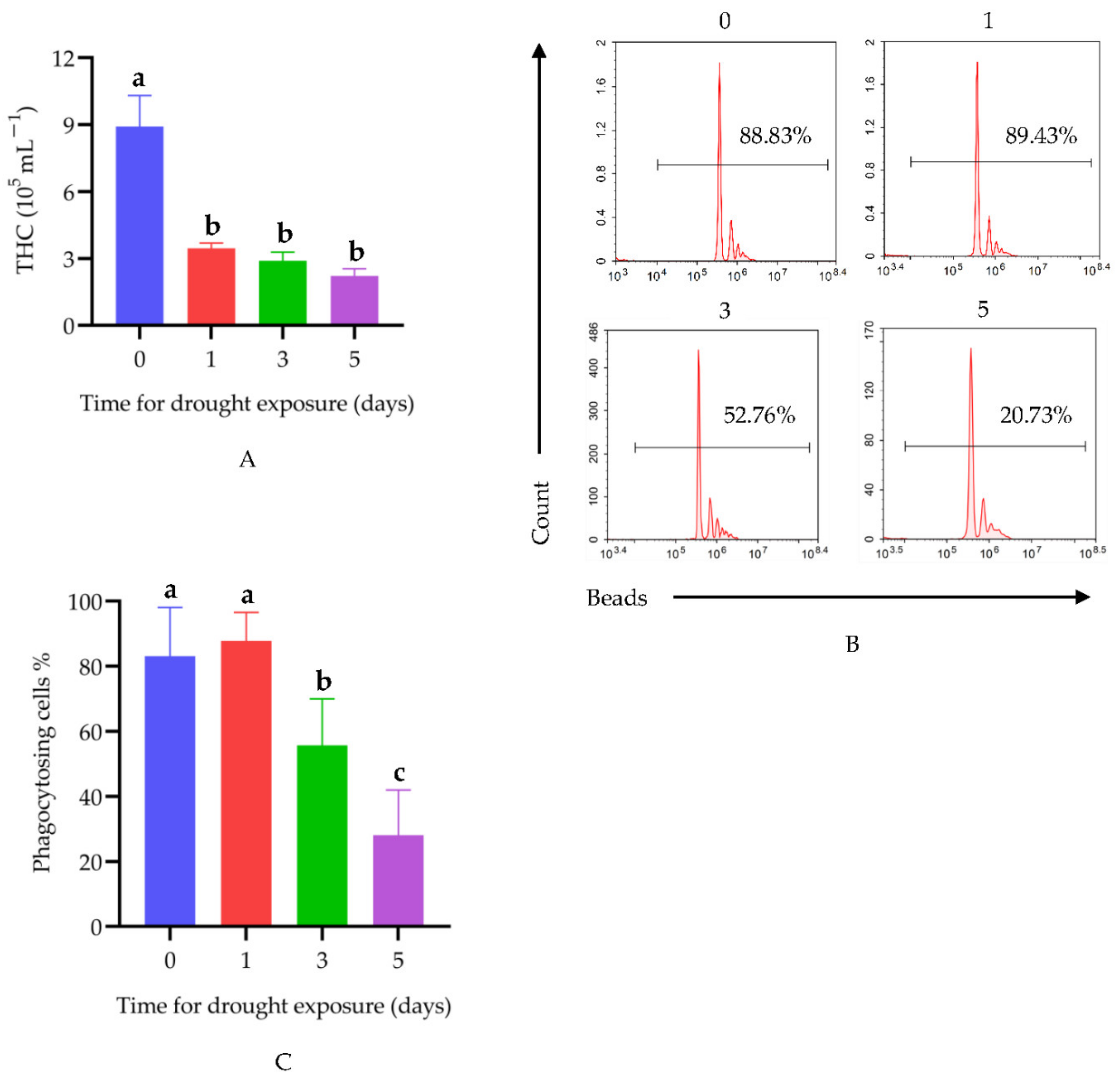
3.1. Drought Suppresses Immune Function
3.2. Drought Promotes Overproduction of ROS in Hepatopancreas
3.3. Drought Induces Histological Disorder in Hepatopancreas
3.4. Drought Induces Metabolic Changes in Hepatopancreas
3.4.1. Quality Control (QC) Analysis
3.4.2. Metabolite Profile of Hepatopancreas
3.4.3. Identification of Potential Biomarkers and Analysis of Related Metabolic Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.; Knowler, D.J.; Leveque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. Camb. Philos. Soc. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Harrison, I.; Abell, R.; Darwall, W.; Thieme, M.L.; Tickner, D.; Timboe, I. The freshwater biodiversity crisis. Science 2018, 362, 1369. [Google Scholar] [CrossRef] [PubMed]
- Richman, N.I.; Bohm, M.; Adams, S.B.; Alvarez, F.; Bergey, E.A.; Bunn, J.J.; Burnham, Q.; Cordeiro, J.; Coughran, J.; Crandall, K.A.; et al. Multiple drivers of decline in the global status of freshwater crayfish (Decapoda: Astacidea). Philos. Trans. R. Soc. B 2015, 370, 20140060. [Google Scholar] [CrossRef] [PubMed]
- Kouba, A.; Tikal, J.; Cisar, P.; Vesely, L.; Fort, M.; Priborsky, J.; Patoka, J.; Buric, M. The significance of droughts for hyporheic dwellers: Evidence from freshwater crayfish. Sci. Rep. 2016, 6, 26569. [Google Scholar] [CrossRef]
- Jeremias, G.; Barbosa, J.; Marques, S.M.; Asselman, J.; Goncalves, F.J.M.; Pereira, J.L. Synthesizing the role of epigenetics in the response and adaptation of species to climate change in freshwater ecosystems. Mol. Ecol. 2018, 27, 2790–2806. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Weiperth, A.; Hossain, M.S.; Kubec, J.; Grabicova, K.; Lozek, F.; Vesely, L.; Blaha, M.; Buric, M.; Kouba, A.; et al. The effects of the herbicides terbuthylazine and metazachlor at environmental concentration on the burrowing behaviour of red swamp crayfish. Chemosphere 2021, 270, 128656. [Google Scholar] [CrossRef]
- Guo, W.; Hossain, M.S.; Kubec, J.; Grabicova, K.; Randak, T.; Buric, M.; Kouba, A. Psychoactive compounds at environmental concentration alter burrowing behavior in the freshwater crayfish. Sci. Total Environ. 2020, 711, 135138. [Google Scholar] [CrossRef]
- Matthews, W.J.; Marsh-Matthews, E. Effects of drought on fish across axes of space, time and ecological complexity. Freshw. Biol. 2003, 48, 1232–1253. [Google Scholar] [CrossRef]
- Alkan Uckun, A.; Barim Oz, O. Acute exposure to the fungicide penconazole affects some biochemical parameters in the crayfish (Astacus leptodactylus Eschscholtz, 1823). Environ. Sci. Pollut. Res. Int. 2020, 27, 35626–35637. [Google Scholar] [CrossRef]
- Volpe, M.G.; Ghia, D.; Safari, O.; Paolucci, M. Fast non-destructive assessment of heavy metal presence by ATR-FTIR analysis of crayfish exoskeleton. Environ. Sci. Pollut. Res. Int. 2020, 27, 21021–21031. [Google Scholar] [CrossRef]
- Gherardi, F. Invasive crayfish and freshwater fishes of the world. Rev. Sci. Technol. 2010, 29, 241–254. [Google Scholar] [CrossRef]
- Calderon-Rosete, G.; Gonzalez-Barrios, J.A.; Lara-Lozano, M.; Pina-Leyva, C.; Rodriguez-Sosa, L. Transcriptional Identification of Related Proteins in the Immune System of the Crayfish Procambarus clarkii. High-Throughput 2018, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Bouallegui, Y. A Comprehensive Review on Crustaceans’ Immune System With a Focus on Freshwater Crayfish in Relation to Crayfish Plague Disease. Front. Immunol. 2021, 12, 667787. [Google Scholar] [CrossRef]
- Soderhall, I. Crustacean hematopoiesis. Dev. Comp. Immunol. 2016, 58, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Li, J.; Chen, Y.; Si, Q.; Tian, J.; Jiang, Q.; Yang, J. Exposure time relevance of response to nitrite exposure: Insight from transcriptional responses of immune and antioxidant defense in the crayfish, Procambarus clarkii. Aquat. Toxicol. 2019, 214, 105262. [Google Scholar] [CrossRef]
- Liu, F.; Qu, Y.K.; Geng, C.; Wang, A.M.; Zhang, J.H.; Chen, K.J.; Liu, B.; Tian, H.Y.; Yang, W.P.; Yu, Y.B. Effects of hesperidin on the growth performance, antioxidant capacity, immune responses and disease resistance of red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2020, 99, 154–166. [Google Scholar] [CrossRef]
- Guo, K.; Ruan, G.; Fan, W.; Fang, L.; Wang, Q.; Luo, M.; Yi, T. The effect of nitrite and sulfide on the antioxidant capacity and microbial composition of the intestines of red swamp crayfish, Procambarus clarkii. Fish Shellfish Immunol. 2020, 96, 290–296. [Google Scholar] [CrossRef]
- Hossain, M.M.; Huang, H.; Yuan, Y.; Wan, T.; Jiang, C.; Dai, Z.; Xiong, S.; Cao, M.; Tu, S. Silicone stressed response of crayfish (Procambarus clarkii) in antioxidant enzyme activity and related gene expression. Environ. Pollut. 2021, 274, 115836. [Google Scholar] [CrossRef]
- Hong, Y.; Huang, Y.; Yan, G.; Yin, H.; Huang, Z. DNA damage, immunotoxicity, and neurotoxicity induced by deltamethrin on the freshwater crayfish, Procambarus clarkii. Environ. Toxicol. 2020, 36, 16–23. [Google Scholar] [CrossRef]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.P.; Fleury, M.J.J. Microbial antioxidant defense enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef]
- Judge, A.; Dodd, M.S. Metabolism. Essays Biochem. 2020, 64, 607–647. [Google Scholar] [CrossRef]
- Gallego, O.; Betts, M.J.; Gvozdenovic-Jeremic, J.; Maeda, K.; Matetzki, C.; Aguilar-Gurrieri, C.; Beltran-Alvarez, P.; Bonn, S.; Fernandez-Tornero, C.; Jensen, L.J.; et al. A systematic screen for protein-lipid interactions in Saccharomyces cerevisiae. Mol. Syst. Biol. 2010, 6, 430. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, X.; Zhang, J.; Luo, Y.; Xu, B.; Ling, S.; Zhang, Y.; Li, W.; Yao, X. Activation of aryl hydrocarbon receptor in Langerhans cells by a microbial metabolite of tryptophan negatively regulates skin inflammation. J. Dermatol. Sci. 2020, 100, 192–200. [Google Scholar] [CrossRef]
- Li, X.; Gianoulis, T.A.; Yip, K.Y.; Gerstein, M.; Snyder, M. Extensive in vivo metabolite-protein interactions revealed by large-scale systematic analyses. Cell 2010, 143, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Dang, M.; Li, Y.; Song, J. ATP biphasically modulates LLPS of SARS-CoV-2 nucleocapsid protein and specifically binds its RNA-binding domain. Biochem. Biophys. Res. Commun. 2021, 541, 50–55. [Google Scholar] [CrossRef]
- Dieterich, I.A.; Lawton, A.J.; Peng, Y.; Yu, Q.; Rhoads, T.W.; Overmyer, K.A.; Cui, Y.; Armstrong, E.A.; Howell, P.R.; Burhans, M.S.; et al. Acetyl-CoA flux regulates the proteome and acetyl-proteome to maintain intracellular metabolic crosstalk. Nat. Commun. 2019, 10, 3929. [Google Scholar] [CrossRef]
- Nakahata, Y.; Kaluzova, M.; Grimaldi, B.; Sahar, S.; Hirayama, J.; Chen, D.; Guarente, L.P.; Sassone-Corsi, P. The NAD+-dependent deacetylase SIRT1 modulates CLOCK-mediated chromatin remodeling and circadian control. Cell 2008, 134, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Kim, A.; Lee, W.; Kim, S.; Lee, S.; Yoon, D.; Bae, J.S.; Park, C.I.; Kim, S. Vibrio harveyi Infection Significantly Alters Amino Acid and Carbohydrate Metabolism in Whiteleg Shrimp, Litopenaeus vannamei. Metabolites 2020, 10, 265. [Google Scholar] [CrossRef]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies-Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.W.; Wang, Y.; Wang, X.W.; Wang, L.; Mu, Y.; Wang, J.X. A C-type lectin with an immunoglobulin-like domain promotes phagocytosis of hemocytes in crayfish Procambarus clarkii. Sci. Rep. 2016, 6, 29924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Li, S.; Yu, Y.; Sun, M.; Xiang, J.; Li, F. Effects of ammonia stress on the hemocytes of the Pacific white shrimp Litopenaeus vannamei. Chemosphere 2020, 239, 124759. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Chang, X.; Xu, L.; Yang, F. Different roles of crayfish hemocytes in the uptake of foreign particles. Fish Shellfish Immunol. 2018, 77, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Loker, E.S.; Adema, C.M.; Zhang, S.M.; Kepler, T.B. Invertebrate immune systems--not homogeneous, not simple, not well understood. Immunol. Rev. 2004, 198, 10–24. [Google Scholar] [CrossRef]
- He, Z.; Mao, F.; Lin, Y.; Li, J.; Zhang, X.; Zhang, Y.; Xiang, Z.; Noor, Z.; Zhang, Y.; Yu, Z. Molecular characteristics of AMPK and its role in regulating the phagocytosis of oyster hemocytes. Fish Shellfish Immunol. 2019, 93, 416–427. [Google Scholar] [CrossRef]
- Gao, X.; Liu, X.; Song, X.; Teng, P.; Ji, H.; Peng, L.; Qiu, Y.; Guo, D.; Jiang, S. Effect of maduramicin on crayfish (Procambius clarkii): Hematological parameters, oxidative stress, histopathological changes and stress response. Ecotoxicol. Environ. Saf. 2021, 211, 111896. [Google Scholar] [CrossRef]
- Ainerua, M.O.; Tinwell, J.; Murphy, R.; Galli, G.L.J.; van Dongen, B.E.; White, K.N.; Shiels, H.A. Prolonged phenanthrene exposure reduces cardiac function but fails to mount a significant oxidative stress response in the signal crayfish (Pacifastacus leniusculus). Chemosphere 2021, 268, 129297. [Google Scholar] [CrossRef]
- Capanni, F.; Greco, S.; Tomasi, N.; Giulianini, P.G.; Manfrin, C. Orally administered nano-polystyrene caused vitellogenin alteration and oxidative stress in the red swamp crayfish (Procambarus clarkii). Sci. Total Environ. 2021, 791, 147984. [Google Scholar] [CrossRef]
- Yang, Y.; Bazhin, A.V.; Werner, J.; Karakhanova, S. Reactive oxygen species in the immune system. Int. Rev. Immunol. 2013, 32, 249–270. [Google Scholar] [CrossRef]
- Al-Shehri, S.S. Reactive oxygen and nitrogen species and innate immune response. Biochimie 2021, 181, 52–64. [Google Scholar] [CrossRef]
- Rodrigues, S.; Antunes, S.C.; Nunes, B.; Correia, A.T. Histopathological effects of the antibiotic erythromycin on the freshwater fish species Oncorhynchus mykiss. Ecotoxicol. Environ. Saf. 2019, 181, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Kholodkevich, S.; Sharov, A.; Feng, Y.; Ren, N.; Sun, K. Microcystin-LR-induced changes of hepatopancreatic transcriptome, intestinal microbiota, and histopathology of freshwater crayfish (Procambarus clarkii). Sci. Total Environ. 2020, 711, 134549. [Google Scholar] [CrossRef]
- Chabera, J.; Stara, A.; Kubec, J.; Buric, M.; Zuskova, E.; Kouba, A.; Velisek, J. The effect of chronic exposure to chloridazon and its degradation product chloridazon-desphenyl on signal crayfish Pacifastacus leniusculus. Ecotoxicol. Environ. Saf. 2021, 208, 111645. [Google Scholar] [CrossRef]
- Lutnicka, H.; Bojarski, B.; Witeska, M.; Tombarkiewicz, B.; Formicki, G. Exposure to herbicide linuron results in alterations in hematological profile and stress biomarkers of common carp (Cyprinus carpio). Ecotoxicology 2019, 28, 69–75. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing amino acids: An overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [CrossRef] [Green Version]
- Hrncic, D.; Rasic-Markovic, A.; Macut, D.; Mladenovic, D.; Susic, V.; Djuric, D.; Stanojlovic, O. Sulfur-Containing Amino Acids in Seizures: Current State of the Art. Curr. Med. Chem. 2018, 25, 378–390. [Google Scholar] [CrossRef]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Stipanuk, M.H.; Ueki, I. Dealing with methionine/homocysteine sulfur: Cysteine metabolism to taurine and inorganic sulfur. J. Inherit. Metab. Dis. 2011, 34, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Dorman, D.C.; Moulin, F.J.; McManus, B.E.; Mahle, K.C.; James, R.A.; Struve, M.F. Cytochrome oxidase inhibition induced by acute hydrogen sulfide inhalation: Correlation with tissue sulfide concentrations in the rat brain, liver, lung, and nasal epithelium. Toxicol. Sci. 2002, 65, 18–25. [Google Scholar] [CrossRef]
- Schwarz, J.; Konjik, V.; Jankowitsch, F.; Sandhoff, R.; Mack, M. Identification of the Key Enzyme of Roseoflavin Biosynthesis. Angew. Chem. Int. Ed. Engl. 2016, 55, 6103–6106. [Google Scholar] [CrossRef]
- Lee, E.R.; Blount, K.F.; Breaker, R.R. Roseoflavin is a natural antibacterial compound that binds to FMN riboswitches and regulates gene expression. RNA Biol. 2009, 6, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Mansjo, M.; Johansson, J. The riboflavin analog roseoflavin targets an FMN-riboswitch and blocks Listeria monocytogenes growth, but also stimulates virulence gene-expression and infection. RNA Biol. 2011, 8, 674–680. [Google Scholar] [CrossRef] [Green Version]
- Ott, E.; Stolz, J.; Lehmann, M.; Mack, M. The RFN riboswitch of Bacillus subtilis is a target for the antibiotic roseoflavin produced by Streptomyces davawensis. RNA Biol. 2009, 6, 276–280. [Google Scholar] [CrossRef] [Green Version]
- Dawlaty, J.; Zhang, X.; Fischbach, M.A.; Clardy, J. Dapdiamides, tripeptide antibiotics formed by unconventional amide ligases. J. Nat. Prod. 2010, 73, 441–446. [Google Scholar] [CrossRef]
- Kuroishi, T. Regulation of immunological and inflammatory functions by biotin. Can. J. Physiol. Pharmacol. 2015, 93, 1091–1096. [Google Scholar] [CrossRef]
- Mall, G.K.; Chew, Y.C.; Zempleni, J. Biotin requirements are lower in human Jurkat lymphoid cells but homeostatic mechanisms are similar to those of HepG2 liver cells. J. Nutr. 2010, 140, 1086–1092. [Google Scholar] [CrossRef] [Green Version]
- Crisp, S.E.; Griffin, J.B.; White, B.R.; Toombs, C.F.; Camporeale, G.; Said, H.M.; Zempleni, J. Biotin supply affects rates of cell proliferation, biotinylation of carboxylases and histones, and expression of the gene encoding the sodium-dependent multivitamin transporter in JAr choriocarcinoma cells. Eur. J. Nutr. 2004, 43, 23–31. [Google Scholar] [CrossRef]
- Agrawal, S.; Agrawal, A.; Said, H.M. Biotin deficiency enhances the inflammatory response of human dendritic cells. Am. J. Physiol. Cell Physiol. 2016, 311, C386–C391. [Google Scholar] [CrossRef]
- Skupsky, J.; Sabui, S.; Hwang, M.; Nakasaki, M.; Cahalan, M.D.; Said, H.M. Biotin Supplementation Ameliorates Murine Colitis by Preventing NF-kappaB Activation. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Attia, H.; Albuhayri, S.; Alaraidh, S.; Alotaibi, A.; Yacoub, H.; Mohamad, R.; Al-Amin, M. Biotin, coenzyme Q10, and their combination ameliorate aluminium chloride-induced Alzheimer’s disease via attenuating neuroinflammation and improving brain insulin signaling. J. Biochem. Mol. Toxicol. 2020. [Google Scholar] [CrossRef]
- He, P.; Jiang, W.D.; Liu, X.A.; Feng, L.; Wu, P.; Liu, Y.; Jiang, J.; Tan, B.P.; Yang, Q.H.; Kuang, S.Y.; et al. Dietary biotin deficiency decreased growth performance and impaired the immune function of the head kidney, spleen and skin in on-growing grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2020, 97, 216–234. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.R.; Masefield, S.; Hunt, R.; Wood, M.J.; Hart, A.G.; Hallam, J.; Griffiths, S.W.; Cable, J. Terrestrial emigration behaviour of two invasive crayfish species. Behav. Process. 2019, 167, 103917. [Google Scholar] [CrossRef] [PubMed]
- Dragicevic, P.; Grbin, D.; Maguire, I.; Blazevic, S.A.; Abramovic, L.; Tarandek, A.; Hudina, S. Immune Response in Crayfish Is Species-Specific and Exhibits Changes along Invasion Range of a Successful Invader. Biology 2021, 10, 1102. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | Flow Rate (mL/min) | Negative Ion Mode | Positive Ion Mode | ||
|---|---|---|---|---|---|
| A (%) | B (%) | C (%) | D (%) | ||
| 0 | 0.25 | 98 | 2 | 98 | 2 |
| 1 | 0.25 | 98 | 2 | 98 | 2 |
| 9 | 0.25 | 50 | 50 | 50 | 50 |
| 12 | 0.25 | 2 | 98 | 2 | 98 |
| 13.5 | 0.25 | 2 | 98 | 2 | 98 |
| 14 | 0.25 | 98 | 2 | 98 | 2 |
| 17 | 0.25 | 98 | 2 | 98 | 2 |
| No. | Name | Molecular Formula | Retention Time (min) | Mode (C18) | FC Value | p Value | VIP Value | Main Metabolic Pathways |
|---|---|---|---|---|---|---|---|---|
| 1 | 4-Aminobutyric acid | C4H9NO2 | 1.23 | Pos | −1.85 | 0.02 | 1.47 | cAMP signaling pathway GABAergic synapse Estrogen signaling pathway GnRH secretion |
| 2 | Biotin | C10H16N2O3S | 1.28 | Pos | −1.52 | 0.05 | 1.50 | Biotin metabolism Biosynthesis of cofactors |
| 3 | Dethiobiotin | C10H18N2O3 | 10.52 | Pos | 1.70 | 0.00 | 2.19 | Biotin metabolism Biosynthesis of cofactors |
| 4 | D-O-Phosphoserine | C3H8NO6P | 1.13 | Pos | −2.09 | 0.01 | 1.79 | Aminoacyl-tRNA biosynthesis Biosynthesis of various other secondary metabolites Biosynthesis of various antibiotics |
| 5 | Genistein | C15H10O5 | 13.95 | Pos | 9.82 | 0.00 | 1.94 | Isoflavonoid biosynthesis Biosynthesis of secondary metabolites |
| 6 | Glycine anhydride | C4H6N2O2 | 1.79 | Pos | −1.77 | 0.00 | 1.66 | Pyrimidine metabolism beta-Alanine metabolism Pantothenate and CoA biosynthesis |
| 7 | Guanosine-5’-monophosphate | C10H14N5O8P | 2.30 | Pos | −2.60 | 0.01 | 1.44 | Purine metabolism Biosynthesis of alkaloids derived from histidine and purine Olfactory transduction |
| 8 | L-Glutathione oxidized | C20H32N6O12S2 | 3.57 | Pos | −2.44 | 0.01 | 1.51 | Glutathione metabolism Biosynthesis of cofactors |
| 9 | Nicotinamide | C6H6N2O | 2.21 | Pos | −4.05 | 0.00 | 2.21 | Nicotinate and nicotinamide metabolism Metabolic pathways Biosynthesis of cofactors |
| 10 | Ophthalmic acid | C11H19N3O6 | 4.56 | Pos | −1.86 | 0.01 | 1.83 | Cysteine and methionine metabolism Metabolic pathways |
| 11 | (R)-Prunasin | C14H17NO6 | 3.68 | Pos | −5.32 | 0.00 | 2.26 | Metabolic pathways Biosynthesis of secondary metabolites |
| 12 | S-Adenosylhomocysteine | C14H20N6O5S | 3.78 | Pos | −1.52 | 0.02 | 1.50 | Cysteine and methionine metabolism Biosynthesis of cofactors” |
| 13 | S-Glutathionyl-L-cysteine | C13H22N4O8S2 | 1.25 | Pos | −1.65 | 0.03 | 1.55 | Cysteine and methionine metabolism Metabolic pathways |
| 14 | Tyramine | C8H11NO | 4.79 | Pos | −2.10 | 0.01 | 1.52 | Isoquinoline alkaloid biosynthesis Biosynthesis of alkaloids derived from shikimate pathway Biosynthesis of secondary metabolites |
| 15 | Citric acid | C6H8O7 | 1.21 | Neg | −2.90 | 0.00 | 2.33 | Citrate cycle (TCA cycle) Biosynthesis of phenylpropanoids Biosynthesis of alkaloids derived from shikimate pathway |
| 16 | γ-Glutamyltyramine | C13H18N2O4 | 4.15 | Neg | 1.60 | 0.03 | 1.70 | Metabolic pathways Biosynthesis of cofactors |
| 17 | 5-Hydroxy-L-tryptophan | C11H12N2O3 | 4.23 | Neg | −2.93 | 0.01 | 2.03 | Biosynthesis of alkaloids derived from shikimate pathway Serotonergic synapse |
| 18 | Isonocardicin A | C23H24N4O9 | 6.24 | Neg | −2.27 | 0.03 | 1.75 | Monobactam biosynthesis Biosynthesis of secondary metabolites |
| 19 | Misonidazole | C7H11N3O4 | 1.20 | Neg | −1.75 | 0.02 | 1.75 | Biosynthesis of various antibiotics Biosynthesis of secondary metabolites |
| 20 | β-N-Acetylglucosamine | C8H15NO6 | 1.18 | Neg | −1.52 | 0.03 | 1.75 | Amino sugar and nucleotide sugar metabolism Phosphotransferase system (PTS) |
| 21 | Pantothenic acid | C9H17NO5 | 5.96 | Neg | −1.66 | 0.01 | 2.03 | Pantothenate and CoA biosynthesis Biosynthesis of secondary metabolites Biosynthesis of cofactors |
| 22 | S-(5-Deoxy-D-ribos-5-yl)-L-homocysteine | C9H17NO6S | 1.21 | Neg | −1.68 | 0.01 | 1.85 | Cysteine and methionine metabolism Metabolic pathways |
| 23 | S-Glutathionyl-L-cysteine | C13H22N4O8S2 | 1.18 | Neg | −1.81 | 0.01 | 2.02 | Cysteine and methionine metabolism Metabolic pathways |
| 24 | UDP-2-acetamido-2,6-dideoxy-beta-L-talose | C17H27N3O16P2 | 2.01 | Neg | −2.48 | 0.03 | 1.55 | O-Antigen nucleotide sugar biosynthesis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Bai, X.; Li, Y.; Li, J.; Meng, Y.; Xu, Z.; Tang, J.; Lu, Y.; Huang, Y. Changes in the Immunity, Histopathology, and Metabolism of Crayfish (Procambarus clarkii) in Response to Drought. Animals 2022, 12, 890. https://doi.org/10.3390/ani12070890
Xu H, Bai X, Li Y, Li J, Meng Y, Xu Z, Tang J, Lu Y, Huang Y. Changes in the Immunity, Histopathology, and Metabolism of Crayfish (Procambarus clarkii) in Response to Drought. Animals. 2022; 12(7):890. https://doi.org/10.3390/ani12070890
Chicago/Turabian StyleXu, Hui, Xuexia Bai, Yu Li, Jiajia Li, Yong Meng, Zhiqiang Xu, Jianqing Tang, Yan Lu, and Yahong Huang. 2022. "Changes in the Immunity, Histopathology, and Metabolism of Crayfish (Procambarus clarkii) in Response to Drought" Animals 12, no. 7: 890. https://doi.org/10.3390/ani12070890