
Ethnobotany, Phytochemistry, and Biological Activities of the Genus Cordyline

 , , ,
, , ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Botany and Taxonomy
3. Traditional Uses of the Genus Cordyline
4. Other Important Uses of Plants of the Genus Cordyline
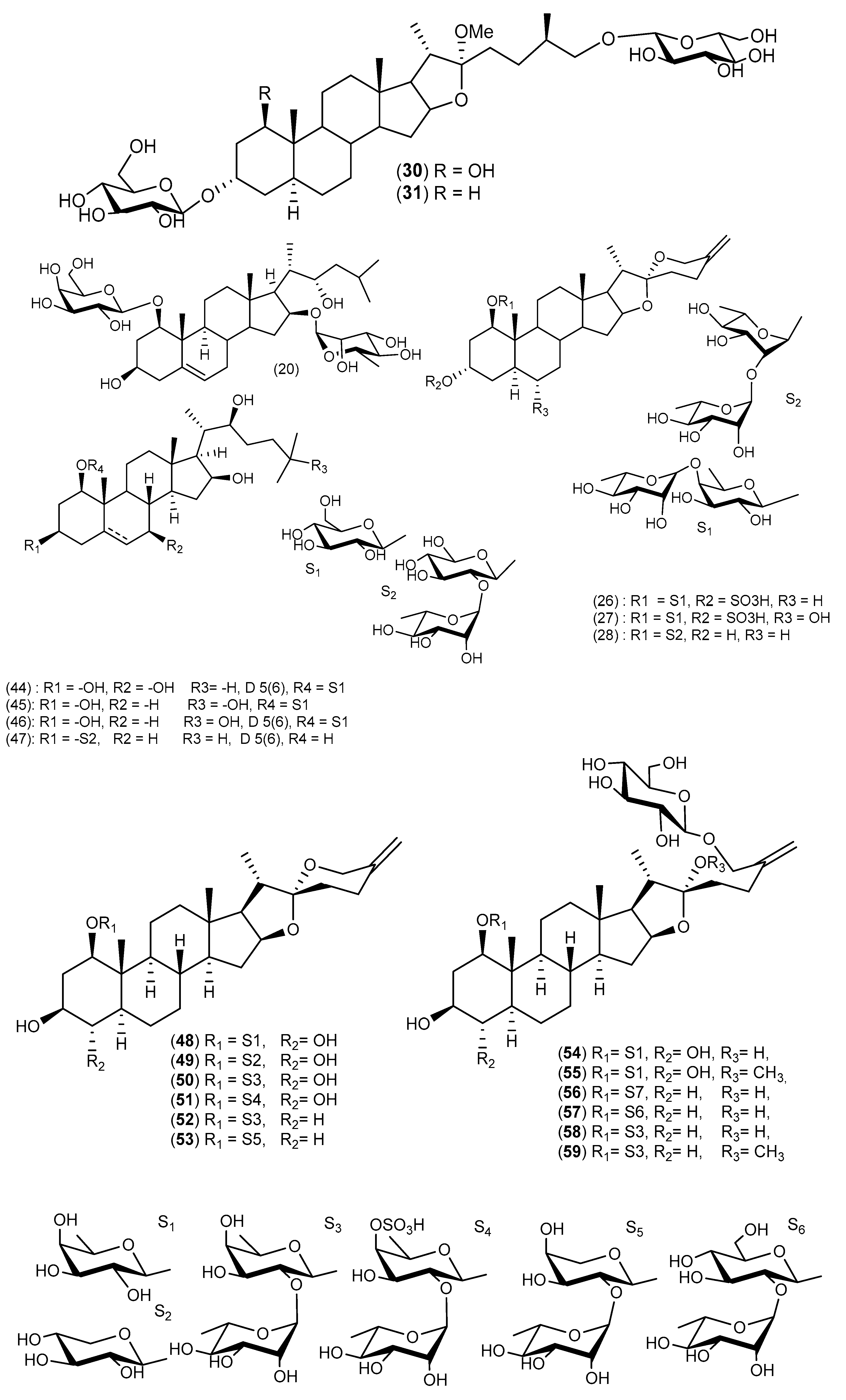
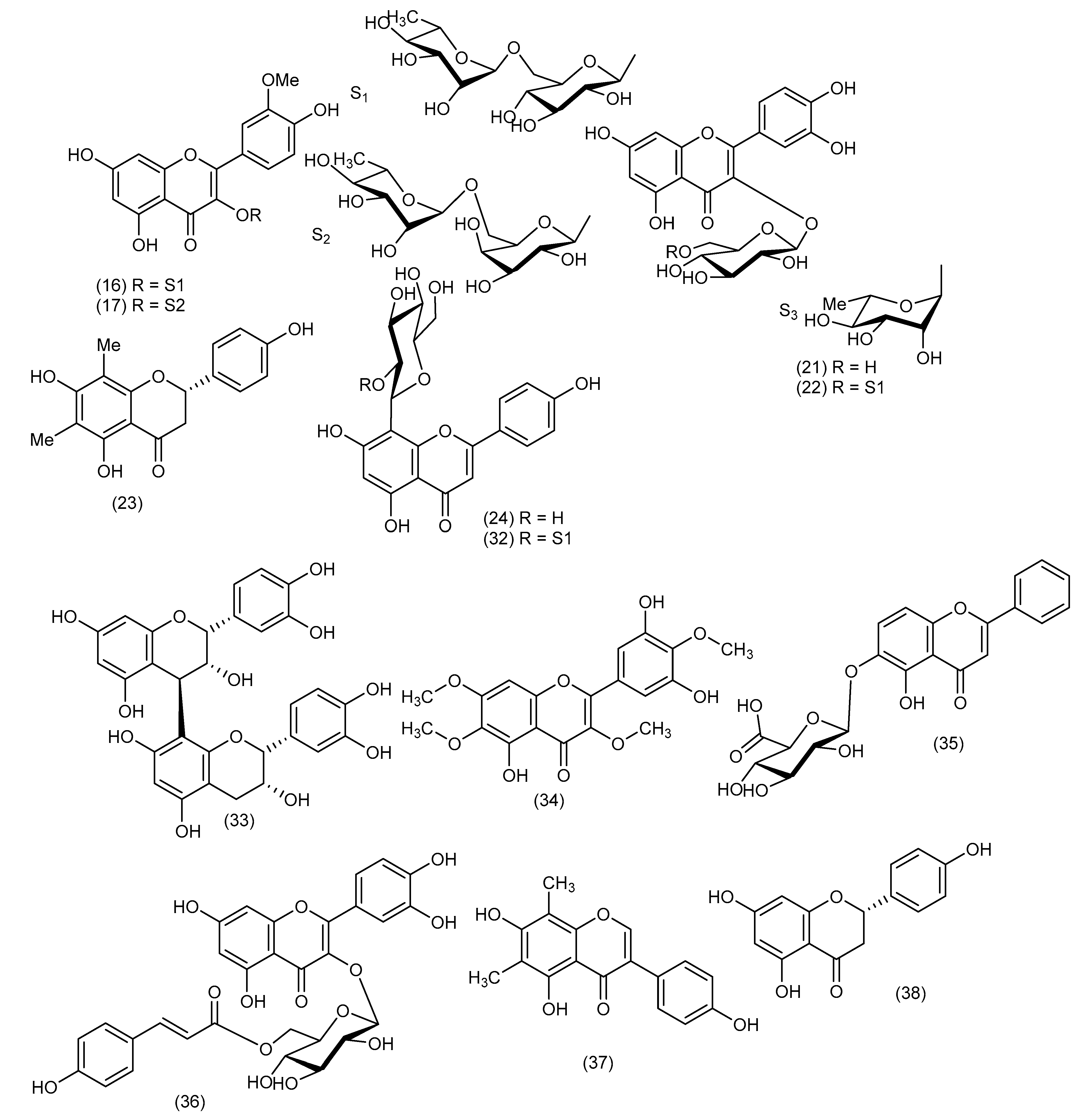
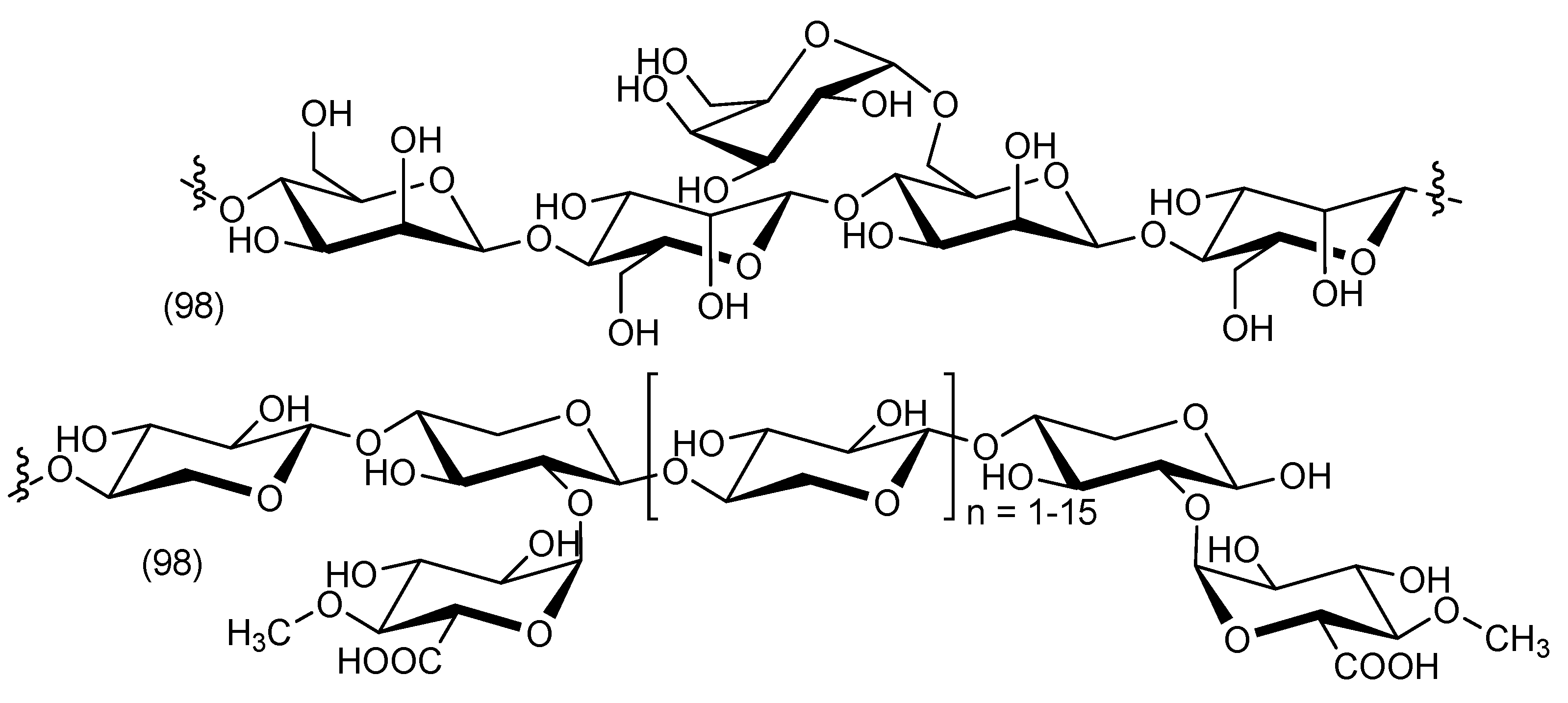
5. Chemical Constituents
5.1. Cordyline australis
5.2. Cordyline fruticosa (Synonym: Cordyline terminalis)
5.3. Cordyline manners-suttoniae
5.4. Cordyline stricta
5.5. Cordyline rubra
5.6. Cordyline cannifolia
5.7. Cordyline dracaenoides
5.8. Cordyline indivisa
6. Biological Activities
6.1. Antimicrobial Activity
6.2. Antioxidant Activity
6.3. Antiproliferative Activity
6.4. Hypolipidemic Potential, Antiparasitic Activity, Nephrotoxicity and Hepatotoxicity Preventions
7. Toxicology
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Porter, R.J.; Medrum, B.S. Antiseizure drug. In Basic & Clinical Pharmacology, 12th ed.; Katzung, B.G., Masters, S.B., Trevor, A.J., Eds.; Prentice Hall International (UK) Ltd: London, UK, 1996; pp. 331–342. [Google Scholar]
- Muazu, J.; Kaita, A.H. A review of traditional plants used in the treatment of epilepsy amongst the Hausa/Fulani tribes of northern Nigeria. Afr. J. Tradit. Complement. Altern. Med. 2008, 5, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–355. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Pezzuto, J.M. Natural products as a vital source for the discovery of cancer chemotherapeutic and chemopreventive agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Sofi, M.S.; Sateesh, M.K.; Bashir, M.; Ganie, M.A.; Nabi, S. Chemopreventive and anti-breast cancer activity of compounds isolated from leaves of Abrus precatorius L. Biotech 2018, 8, 371. [Google Scholar] [CrossRef] [PubMed]
- Blunden, G.; Jaffer, A.J.; Jewers, J.K.; Griffin, W.J. Steroidal sapogenins from leaves of Cordyline species. J. Nat. Prod. 1981, 44, 441–447. [Google Scholar] [CrossRef]
- Fouedjou, R.T.; Teponno, R.B.; Quassinti, L.; Bramucci, M.; Petrelli, D.; Vitali, L.A.; Fiorini, D.; Tapondjou, L.A. Steroidal saponins from the leaves of Cordyline frutcosa (L) A. Chev and their cytotoxic and antimicrobial activity. Phytochem. Lett. 2014, 7, 62–68. [Google Scholar] [CrossRef]
- Tran, T.D.; Olsson, M.A.; Choudhury, M.A.; McMillan, D.; Cullen, J.; Parsons, P.J.; Bernhardt, V.P.; Reddell, P.W.; Ogbourne, M.S. Antibacterial 5α-Spirostane saponins from the fruit of Cordyline manners-suttoniae. J. Nat. Prod. 2019, 82, 2809–2817. [Google Scholar] [CrossRef] [PubMed]
- Mimaki, Y.; Kuroda, M.; Akaashi, Y.; Sashida, Y. Steroidal glucosides from leaves of Cordyline stricta. Phytochemistry 1997, 45, 1229–1234. [Google Scholar] [CrossRef]
- Korkashvili, T. Steroidal glycosides of Cordyline australis. Master’s Thesis, Philosophy in Chemistry, University of Waikato, Hamilton, New Zealand, 2006. Available online: http://hdl.handle.net/10289/2237 (accessed on 8 May 2022).
- Ponou, K.B.; Teponno, B.R.; Tapondjou, A.L.; Lacaille-Dubois, M.; Quassinti, L.; Bramussi, M.; Barboni, L. Steroidal saponins from the aerial parts of Cordyline fruticosa L. Var strawberries. Fitoterapia 2019, 134, 454–458. [Google Scholar] [CrossRef]
- Blunden, G.; Sitton, D.; Beach, S.; Turne, C.H. Australigenin, A new steroidal sapogenin from Cordyline australis fruits. J. Nat. Prod. 1984, 47, 266–269. [Google Scholar] [CrossRef]
- Chunduri, J.R.; Shah, H.R. FTIR phytochemical fingerprinting and antioxidant analysis of selected indoor non-flowering indoor plants and their industrial importance. Int. J. Curr. Pharm. Res. 2016, 8, 37–43. [Google Scholar] [CrossRef]
- Elfita, M.; Muharni; Mardiyanto; Fitrya. Chemical compounds from the antibacterial active fraction of Cordyline fruticosa (L). Conf. Series: Earth Environ. Sci. 2021, 709, 012048. [Google Scholar] [CrossRef]
- Encyclopedia Britannica. Plant-Nomenclature-Taxonomy. Available online: https://www.britannica.com/biography/Michel-Adanson (accessed on 23 March 2022).
- Mckenzie, E.H.C.; Buchanan, P.K.; Johnston, P.R. Checklist of fungi on cabbage trees (Cordyline spp.) and New Zealand flaxes (Phormium spp.) in New Zealand. N. Z. J. Bot. 2005, 43, 119–139. [Google Scholar] [CrossRef]
- Harris, W.; Ross, E.; Beever, E.R.; Heenan, B.P. Phenotypic variation of leaves of and stems of wild stands of Cordyline autralis New Zealand J. New Zealand J. Bot. 1998, 36, 593–604. [Google Scholar] [CrossRef]
- Ehrlich, C. The Ethnobotany of Cordyline fruticosa (L.) A. Chev.: The “Hawaiian Ti plant”. Ph.D. Dissertation, State University at Buffalo, Buffalo, NY, USA, 1999. [Google Scholar]
- Khan, S.; Naz, S.; Saeed, B. In vitro production of Cordyline terminalis for commercialization. Pak. J. Bot. 2004, 36, 757–761. [Google Scholar]
- Kattoor, C.J. Micropropagation Studies in Dracaena and Cordyline. Master’s Thesis, University of Agricultural Sciences, Dharwad, India, 2010. [Google Scholar]
- Wilson, A. Flora of Australia: Oceanic Islands, 49; Australian Government Publishing Service: Canberra, Australia, 1994; ISBN -100644293853. [Google Scholar]
- Conran, J.G. Lomandraceae. The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Volume 3: Flowering Plants Monocotyledons: Lilianae (Except Orchidaceae); Springer: Berlin/Heidelberg, Germany, 1998; pp. 354–365. ISBN 103642083773. [Google Scholar]
- World Checklist of Selected Plant Families (WCSP). Available online: https://wcsp.science.kew.org/ (accessed on 1 November 2018).
- Chifundera, K. Livestock diseases and the traditional medicine in the Bushi area. Kivu province, Democratic Republic of Congo. Afr. Study Monogr. 1998, 19, 13–33. [Google Scholar] [CrossRef]
- Lense, O. The wild plants used as traditional medicines by indigenous people of Manokwari, West Papua. Biodiversitas 2012, 13, 98–106. [Google Scholar] [CrossRef]
- Calixto, J.B.; Lima, T.C.M.; Morato, G.S.; Nicolau, M.; Takahashi, R.N.; Valle, R.M.R.; Schmidt, C.C.; Yunes, R.A. Chemical and pharmacological analysis of the crude aqueous/alcoholic extract from Cordyline dracaenoides. Phytother. Res. 1990, 4, 167. [Google Scholar] [CrossRef]
- Melzer, M.; Sugano, J.; Uchida, J.; Kawate, M.; Borth, W.; Hu, J. Partial characterization of a novel emara-like virus from Cordyline fruticosa (L.) with ti ringspot disease. In Proceedings of the 2014 APS-CPS Joint Meeting, Minneapolis, MA, USA, 9–13 August 2014. [Google Scholar] [CrossRef]
- Hinkle, A.E. Population structure of pacific Cordyline fruticosa (Laxmanniaceae) with implications for human settlement of Polynesia. Am. J. Bot. 2007, 94, 828–839. [Google Scholar] [CrossRef]
- Whistler, W.A. Traditional and herbal medicine in the Cook Islands. J. Ethnopharm. 1985, 13, 239–280. [Google Scholar] [CrossRef]
- Buttner, R. Mansfeld’s Encyclopedia of Agricultural and Horticultural Crops; Hanelt, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Dalimartha, S. Atlas Tumbuhan Obat Indonesia; Puspa Swara: Jakarta, Indonesia, 2007; Volume 4, ISBN 979323573X. [Google Scholar]
- Nugent, J. Permaculture Plants: Agaves and Cacti, 2nd ed.; Sustainable Agriculture Research Institute: College Park, MD, USA, 2006. [Google Scholar]
- Nombo, P.; Leach, J. Reite Plants: An Ethnobotanical Study in Tok Pisin and English; ANU E Press: Canberra, Australia, 2010. [Google Scholar]
- Kulip, J. An ethnobotanical survey of medicinal and other useful plants of Muruts in Sabah, Malaysia. Telopea 2003, 10, 81–98. [Google Scholar] [CrossRef]
- Kent, H.W. Treasury of Hawaiian Words in One Hundred and One Categories. The Masonic Public Library of Hawaii; University of Hawaii Press: Honolulu, HI, USA, 1995; 504p. [Google Scholar]
- Ray, T.; Saha, P.; Roy, S.C. Commercial production of Cordyline terminalis (L) Kunth. From shoot apex meristem and assessment for genetic stability of somaclones by isozyme markers. Sci. Hortic. 2006, 108, 289–294. [Google Scholar] [CrossRef]
- Brasch, D.J.; Fankhauser, B.L.; McDonald, A.G. A study of the glucofructofuranon from the New Zealand cabbage tree Cordyline australis. Carbohydr. Res. 1988, 180, 315–324. [Google Scholar] [CrossRef]
- Simpson, P. Dancing Leaves. The Story of New Zealand’s Cabbage Tree, TıKouka; Christchurch, Canterbury University Press: Canterbury, New Zealand, 2000. [Google Scholar]
- Aufenanger, H. The Cordyline plant in the central highlands of new Guinea. Anthropos 1961, 56, 393–408. [Google Scholar]
- Merlin, M. The traditional geographical range and ethnobotanical diversity of Cordyline fruticosa (L.) Chevalier. Ethnobotany 1989, 1, 25–39. [Google Scholar]
- Whistler, W.A. Polynesian Herbal Medicine. Lawa’i, Kaua’i, Hawai’i; National Tropical Botanical Garden: Hong Kong, China, 1992. [Google Scholar]
- Elbert, L.L.J.; Roger, G.S. Common Forest Trees of Hawaii (Native and Introduced); Agriculture Hanbook No. 679; US Department of Agriculture, Forest Service: Washington, DC, USA, 1989. [Google Scholar]
- Lim, T.K. Edible Medicinal and Non-medicinal Plants. Volume 12: Modified Stems, Roots, Bulbs; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar] [CrossRef]
- Duncalf, I. Conventional propagation of Cordyline australis. Act. Hortic. 2014, 63–66. [Google Scholar] [CrossRef]
- Simpson, P. Sudden decline in cabbage trees (Cordyline australis). Sci. Res. Intern. Rep. 1991, 107, 1–19. [Google Scholar]
- Marker, R.E.; Wagner, R.B.; Ulshafer, P.R.; Wittbecker, E.L.; Goldsmith, D.P.J.; Ruof, C.H. Sterols. CLVII. Sapogenins. 69. Isolation and structures of thirteen new steroidal sapogenins. New sources for known sapogenins. J. Am. Chem. Soc. 1943, 65, 1199–1209. [Google Scholar] [CrossRef]
- Wall, M.E.; Eddy, C.R.; Willaman, J.J.; Correll, D.S.; Schubert, B.G.; Gentry, H.S. Steroidal sapogenins. XII. Survey of plants for steroidal sapogenins and other constituents. J. Am. Pharm. Assoc. 1954, 43, 503–505. [Google Scholar] [CrossRef]
- Jewers, A.H.K.; Manchanda, J.D.; Nagle, M.J. Cordylagenin, a new steroidal saponin diol from Cordyline cannifolia and Cordyline stricta. Tetrahedron Lett. 1974, 16, 1475–1478. [Google Scholar] [CrossRef]
- Blunden, G.; Jaffer, J.A.; Jewers, K.; Robinson, J.M. Configurational assignments to spirostan-3-ols by mass spectrometry. Steroids 1980, 36, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Griffin, W.J.; Maunwongyathi, P. A comparison of four species of Cordyline. Planta Med. 1969, 4, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Blunden, G.; Jaffer, J.A.; Jewers, K.; Griffin, W.J. Crabbogenin 1β-hydroxycrabbogenin, strictagenin and pompeygenin, four new steroidal sapogenins from Cordyline stricta leaves. Tetrahedron 1981, 37, 2911–2915. [Google Scholar] [CrossRef]
- Djohan, H.; Widiyantoro, A.; Shofiyani, A. Isolation of flavonoid from Andong leaves (Cordyline fruticosa (L.) A. Chev.) and its activity as complexor of Fe2+. Indones. J. Pure Appl. Chem. 2019, 2, 39–45. [Google Scholar] [CrossRef]
- Raslan, M.; Taher, R.F.; Al-Karmalawy, A.A.; El-Ebeedy, D.; Metwaly, A.G.; Elkateeb, N.M.; Ghanem, A.; Elghaish, R.A.; El Maksoud, A.I.A. Cordyline fruticosa (L.) A. Chev. leaves: Isolation, HPLC/MS profiling and evaluation of nephroprotective and hepatoprotective activities supported by molecular docking. New J. Chem. 2021, 45, 22216–22233. [Google Scholar] [CrossRef]
- Adaku, C.; Skaar, I.; Byamukama, R.; Jordheim, M.; Andersen, Ø.M. Anthocyanin profile and antioxidant property of anti-asthma flowers of Cordyline terminalis (L.) Kunth (Agavaceae). Nat. Prod. Commun. 2020, 15, 1–7. [Google Scholar] [CrossRef]
- Yokosuka, A.; Suzuki, T.; Mimaki, Y. New cholestane glycosides from the l eaves of Cordyline terminalis. Chem. Pharm. Bull. 2012, 60, 275–279. [Google Scholar] [CrossRef]
- Nguyen, D.H.; Mitaine-Offer, A.C.; Miyamoto, T.; Tanaka, C.; Bellaye, P.S.; Collin, B.; Chambin, O.; Lacaille-Dubois, M.A. Steroidal glycosides from the Vietnamese cultivar Cordyline fruticosa “Fairchild red”. Phytochemistry 2021, 192, 112966. [Google Scholar] [CrossRef]
- Yang, M.; Blunden, G.; Patel, A.; Crabb, T.A.; Griffint, W.J. Two furostane sapogenins from cordyline rubra. Phytochemistry 1990, 29, 1332–1334. [Google Scholar] [CrossRef]
- Simmons-Boycea, J.L.; and Tinto, W.F. Steroidal saponins and sapogenins from the Agavaceae family. Nat. Prod. Commun. 2006, 2, 99–114. [Google Scholar] [CrossRef]
- Sidana, J.; Singh, B.; Sharma, O.P. Saponins of Agave: Chemistry and bioactivity. Phytochemistry 2016, 130, 22–46. [Google Scholar] [CrossRef] [PubMed]
- Almaraz-Abarca, N.; Delgado-Alvarado, E.A.; Ávila-Reyes, J.A.; Uribe-Soto, J.N.; González-Valdez, L.S. The phenols of the genus Agave (Agavaceae). J. Biomater. Nanobiotechnol. 2013, 4, 9–16. [Google Scholar] [CrossRef]
- Mimaki, Y.; Kuroda, M.; Akaashi, Y.; Sashida, Y. Steroidal saponins from the leaves of Cordyline stricta. Phytochemistry 1998, 47, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Jewers, K.; Burbage, M.B.; Blunden, G.; Griffin, W.J. Brisbagenin and brisbenone, two new spirostanes from Cordyline species. Steroids 1974, 24, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Griffin, W.; Blunden, G.; Jewers, K.; Burbage, B.M.; Nagler, M.J. Steroidal sapogenins from Cordyline cannifolia leaves. Phytochemistry 1976, 15, 1271–1274. [Google Scholar] [CrossRef]
- Yang, M.; Blunden, G.; Patel, A.; Crabb, T.A.; Griffint, W.J. Rubragenin, chenogenin and wallogenin, steroidal sapogenins from cordyline rubra. Phytochemistry 1989, 28, 3171–3173. [Google Scholar] [CrossRef]
- Sieber, R. Die hemicellulosen von Cordyline indivisa. Phytochemistry 1972, 11, 1433–1441. [Google Scholar] [CrossRef]
- Firoj, A.; Prabir, K.D.; Islam, M.A.; Rahman, K.M.; Rahman, M.M.; Selim, M.S.T. Antibacterial activity of Cordylline terminalis Kunth leaves. J. Med. Sci. 2003, 3, 418–422. [Google Scholar] [CrossRef]
- Prihambodo, T.R.; Nahrowi; Jayanegara, A. Antibacterial activity and phytochemical content of silage juice from tropical herbal leaves. IOP Conf. Ser. Mater. Sci. Eng. 2019, 546, 042032. [Google Scholar] [CrossRef]
- Elfita, E.; Mardiyanto; Fitrya; Larasati, J.E.; Julinar; Widjajanti, H.; Muharni. Antibacterial activity of Cordyline fruticosa leaf extracts and its endophytic fungi extracts. Biodiversitas 2019, 20, 3804–3812. [Google Scholar] [CrossRef]
- Kusuma, I.W.; Sari, N.M.; Murdiyanto; Kuspradini, H. Anticandidal activity of several plants used by Bentian tribe in East Kalimantan, Indonesia. AIP Conf. Proc. 2016, 1755, 040002. [Google Scholar] [CrossRef]
- Reddy, C.; Noor, A.; Sarada, N.C.; Vijayalakshmi, M.A. Antioxidant properties of Cordyline terminalis (L.) Kunth and Myristica fragrans Houtt. Encapsulated separately into casein beads. Curr. Sci. 2011, 101, 416–420. [Google Scholar]
- Sharmila, D.; Kumar, R.S. Antioxidant potential of copper oxide nanoparticles using leaves of Cordyline fruticosa. Drug Invent. Today 2019, 12, 2440–2443. [Google Scholar]
- Banjarnahor, S.D.S.; Artanti, N. Antioxidant properties of flavonoids. Med. J. Indones. 2014, 23, 239–244. [Google Scholar] [CrossRef]
- Tsakem, B.; Tchuenguem, R.T.; Siwe-Noundou, X.; Ponou, K.B.; Dzoyem, J.P.; Teponno, R.B.; Krause, R.W.M.; Tapondjou, A.L. New bioactive flavonoid glycosides with antioxidant activity from the stem bark of Olax subscorpioidea Oliv. Nat. Prod. Res. 2022, 37, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-R.; Zhang, Y.; Jacob, M.R.; Khan, S.I.; Zhang, Y.-J.; Li, X.-C. Antifungal activity of C-27 steroidal saponins. Antimicrob. Agents Chemother. 2006, 50, 1710–1714. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Sánchez, M.L.; Sánchez-Sánchez, L.; Sandoval-Ramírez, J. Steroidal Saponins and Cell Death in Cancer, Chapter 15. In Cell Death—Autophagy, Apoptosis and Necrosis; IntechOpen: London, UK, 2015; pp. 331–352. [Google Scholar] [CrossRef]
- Sobolewska, D.; Galanty, A.; Grabowska, K.; Makowska-Was, J.; Wro’bel-Biedrawa, D.; Podolak, I. Saponins as cytotoxic agents: An update (2010–2018). Part I—Steroidal saponins. Phytochem. Rev 2020, 19, 139–189. [Google Scholar] [CrossRef]
- Zhu, L.; Tan, J.; Wang, B.; Guan, L.; Liu, Y.; Zheng, C. In-vitro antitumor activity and antifungal activity of pennogenin steroidal saponins from Paris Polyphylla var. yunnanensis. Iran. J. Pharm. Res. 2011, 10, 279–286. [Google Scholar] [PubMed]
- Bogoriani, N.W.; Ariati, N.K. The activity of Bali Andong rhizome extract of Cordyline Terminalis Kunth as Hypolipidemia agent in Wistar rats with high-cholesterol diet. Int. J. Pharm. Phytopharmacol. Res. 2018, 8, 75–80. [Google Scholar]
- Bogoriani, N.W.; Suaniti, N.M.; Putra, A.A.B.; Lestari, K.D.P. The activity of Cordyline terminalis’s leaf extract as antidiabetic in obese wistar rats. Int. J. Pharm. Res. Allied Sci. 2019, 8, 206–213. [Google Scholar]
- Dyary, H.O.; Arifah, A.K.; Sharma, R.S.K.; Rasedee, A. Antitrypanosomal and cytotoxic activities of selected medicinal plants and effect of Cordyline terminalis on trypanosomal nuclear and kinetoplast replication. Pak. Vet. J. 2014, 34, 444–448. [Google Scholar]
- Naher, S.; Aziz, A.M.; Akter, I.; Rahman, S.M.; Sajon, S.R.; Mazumder, K. Anti-diarrheal activity and brine shrimp lethality bioassay of methanolic extract of Cordyline fruticosa (L.) A. Chev. Leaves. Clin. Phytosci. 2019, 5, 15. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tematio Fouedjou, R.; Tsakem, B.; Siwe-Noundou, X.; Dongmo Fogang, H.P.; Tiombou Donkia, A.; Kemvoufo Ponou, B.; Poka, M.; Demana, P.H.; Teponno, R.B.; Azefack Tapondjou, L. Ethnobotany, Phytochemistry, and Biological Activities of the Genus Cordyline. Biomolecules 2023, 13, 1783. https://doi.org/10.3390/biom13121783
Tematio Fouedjou R, Tsakem B, Siwe-Noundou X, Dongmo Fogang HP, Tiombou Donkia A, Kemvoufo Ponou B, Poka M, Demana PH, Teponno RB, Azefack Tapondjou L. Ethnobotany, Phytochemistry, and Biological Activities of the Genus Cordyline. Biomolecules. 2023; 13(12):1783. https://doi.org/10.3390/biom13121783
Chicago/Turabian StyleTematio Fouedjou, Romuald, Bienvenu Tsakem, Xavier Siwe-Noundou, Hervet P. Dongmo Fogang, Aphalaine Tiombou Donkia, Beaudelaire Kemvoufo Ponou, Madan Poka, Patrick H. Demana, Rémy B. Teponno, and Léon Azefack Tapondjou. 2023. "Ethnobotany, Phytochemistry, and Biological Activities of the Genus Cordyline" Biomolecules 13, no. 12: 1783. https://doi.org/10.3390/biom13121783