
Unravelling the Helianthus tuberosus L. (Jerusalem Artichoke, Kiku-Imo) Tuber Proteome by Label-Free Quantitative Proteomics

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. MS-Based Proteomics Approach Identifies the Tuber Proteome
2.2. PANTHER-Based Proteome Categorization
3. Discussion
3.1. Selected Differentially Modulated Tuber Proteins with Unique Properties
3.2. Tuber Proteins Potentially Associated with Health and Disease Regulatory Metabolism
3.2.1. Central Carbon Metabolism
3.2.2. Vitamins
3.2.3. Diabetes-Related Proteins
3.2.4. Other Antimicrobial Proteins
4. Materials and Methods
4.1. Specification of Samples and Total Protein Extraction
4.2. Sample Preparation and LC-MS/MS for Proteome Analysis
4.3. Q-Exactive MS Analysis
4.4. LC-MS/MS Data Analysis for Proteome Comparisons
4.5. Mass Spectrometry Protein Data Access
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Liava, V.; Karkanis, A.; Danalatos, N.; Tsiropoulos, N. Cultivation Practices, Adaptability and phytochemical composition of Jerusalem artichoke (Helianthus tuberosus L.): A weed with economic value. Agronomy 2021, 11, 914. [Google Scholar] [CrossRef]
- Stanley, J.A.; Stephan, F.N. Biology and Chemistry of Jerusalem Artichoke: Helianthus tuberosus L., 1st ed.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Ma, X.Y.; Zhang, L.H.; Shao, H.B.; Xu, G.; Zhang, F.; Ni, F.T.; Brestic, M. Jerusalem artichoke (Helianthus tuberosus), a medicinal salt-resistant plant has high adaptability and multiple-use values. J. Med. Plants Res. 2011, 5, 1272–1279. [Google Scholar]
- Fang, Y.R.; Liu, J.A.; Steinberger, Y.; Xie, G.H. Energy use efficiency and economic feasibility of Jerusalem artichoke production on arid and coastal saline lands. Ind. Crops Prod. 2018, 117, 131–139. [Google Scholar] [CrossRef]
- Willaman, J.J. Preparation of inulin products. J. Biol. Chem. 1922, 51, 275–283. [Google Scholar] [CrossRef]
- Yang, L.; He, Q.S.; Corscadden, K.; Udenigwe, C.C. The prospects of Jerusalem artichoke in functional food ingredients and bioenergy production. Biotechnol. Rep. 2015, 5, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Long, X.H.; Shao, H.B.; Liu, L.; Liu, L.P.; Liu, Z.P. Jerusalem artichoke: A sustainable biomass feedstock for biorefinery. Renew. Sustain. Energy Rev. 2016, 54, 1382–1388. [Google Scholar] [CrossRef]
- Kocsis, L.; Liebhard, P.; Praznik, W. Effect of seasonal changes on content and profile of soluble carbohydrates in tubers of different varieties of Jerusalem artichoke (Helianthus tuberosus L.). J. Agric. Food Chem. 2007, 55, 9401–9408. [Google Scholar] [CrossRef]
- Slimestad, R.; Seljaasen, R.; Meijer, K.; Skar, S.L. Norwegian-grown Jerusalem artichoke (Helianthus tuberosus L.): Morphology and content of sugars and fructo-oligosaccharides in stems and tubers. J. Sci. Food Agric. 2010, 90, 956–964. [Google Scholar] [CrossRef]
- Danilcenko, H.; Jariene, E.; Slepetiene, A.; Sawicka, B.; Zaldariene, S. The distribution of bioactive compounds in the tubers of organically grown Jerusalem artichoke (Helianthus tuberosus L.) during the growing period. Acta Sci. Pol. Hortorum Cultus 2017, 16, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Santis, D.D.; Frangipane, M.T. Evaluation of chemical composition and sensory profile in Jerusalem artichoke (Helianthus tuberosus L.) tubers: The effect of clones and cooking conditions. Int. J. Gastron. Food Sci. 2017, 11, 25–30. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Xue, F.; Nan, X.; Wang, H.; Hua, D.; Liu, J.; Yang, L.; Jiang, L.; Xiong, B. Nutritional value, bioactivity, and application potential of Jerusalem artichoke (Helianthus tuberosus L.) as a neotype feed resource. Anim. Nutr. 2020, 6, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Kaszás, L.; Kovács, Z.; Nagy, E.; Elhawat, N.; Abdalla, N.; Domokos-Szabolcsy, E. Jerusalem artichoke (Helianthus tuberosus L.) as a potential chlorophyll source for humans and animals nutrition. Environ. Biodivers. Soil Secur. 2018, 2, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Bogucka, B.; Jankowski, K. Jerusalem artichoke: Quality response to potassium fertilization and irrigation in Poland. Agronomy 2020, 10, 1518. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. Medical importance of Helianthus tuberosus—A review. Indo Am. J. Pharm. Sci. 2018, 5, 2159–2166. [Google Scholar]
- Kleessen, B.; Schwarz, S.; Boehm, A.; Fuhrmann, H.; Richter, A.; Henle, T. Jerusalem artichoke and chicory inulin in bakery products affect faecal microbiota of healthy volunteers. Br. J. Nutr. 2007, 98, 540–549. [Google Scholar] [CrossRef] [Green Version]
- Ramnani, P.; Gaudier, E.; Bingham, M.; Van bruggen, P.; Tuohy, K.M.; Gibson, G.R. Prebiotic effect of fruit and vegetal shots containing Jerusalem artichoke inulin: A human intervention study. Br. J. Nutr. 2010, 104, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Ix, J.H.; Sharma, K. Mechanisms linking obesity, chronic kidney disease, and fatty liver disease: The roles of fetuin-A, adiponectin, and AMPK. J. Am. Soc. Nephrol. 2010, 21, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Dharmalingam, M.; Yamasandhi, P.G. Nsonalcoholic Fatty Liver Disease and Type 2 Diabetes Mellitus. Indian J. Endocrinol. Metab. 2018, 22, 421–428. [Google Scholar] [CrossRef]
- Okada, N.; Kobayashi, S.; Moriyama, K.; Miyataka, K.; Abe, S.; Sato, C.; Kawazoe, K. Helianthus tuberosus (Jerusalem artichoke) tubers improve glucose tolerance and hepatic lipid profile in rats fed a high-fat diet. Asian Pac. J. Trop. Med. 2017, 10, 439–443. [Google Scholar] [CrossRef]
- McMacken, M.; Shah, S. A plant-based diet for the prevention and treatment of type 2 diabetes. J. Geriatr. Cardiol. 2017, 14, 342–354. [Google Scholar]
- Bedekar, A.; Shah, K.; Koffas, M. Natural products for type II diabetes treatment. Adv. Appl. Microbiol. 2010, 71, 21–73. [Google Scholar] [PubMed]
- Yang, D.; Du, X.; Yang, Z.; Liang, Z.; Guo, Z.; Liu, Y. Transcriptomics, proteomics, and metabolomics to reveal mechanisms underlying plant secondary metabolism. Eng. Life Sci. 2014, 14, 456–466. [Google Scholar] [CrossRef]
- Baldini, M.; Danuso, F.; Turi, M.; Vannozzi, G.P. Evaluation of new clones of Jerusalem artichoke (Helianthus tuberosus L.) for inulin and sugar yield from stalks and tubers. Ind. Crops Prod. 2004, 19, 25–40. [Google Scholar] [CrossRef]
- Altan, F.; Unal, R. Focus on proteomics and bioinformatics in translational research and plant research of obesity and diabetes. MOJ Proteom. Bioinform. 2014, 1, 36–41. [Google Scholar]
- Pedrete, T.A.; Hauser-Davis, R.A.; Moreira, J.C. Proteomic characterization of medicinal plants used in the treatment of diabetes. Int. J. Biol. Macromol. 2019, 140, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Min, C.W.; Jung, W.Y.; Park, H.J.; Moon, K.-B.; Ko, H.; Sohn, J.-H.; Jeon, J.-H.; Kim, H.-S.; Gupta, R.; Kim, S.T. Label-free quantitative proteomic analysis determines changes in amino acid and carbohydrate metabolism in three cultivars of Jerusalem artichoke tubers. Plant Biotechnol. Rep. 2019, 13, 111–122. [Google Scholar] [CrossRef]
- Zhang, A.; Han, D.; Wang, Y.; Mu, H.; Zhang, T.; Yan, X.; Pang, Q. Transcriptomic and proteomic feature of salt stress-regulated network in Jerusalem artichoke (Helianthus tuberosus L.) root based on de novo assembly sequencing analysis. Planta 2018, 247, 715–732. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Gupta, A.K. Applications of inulin and oligofructose in health and nutrition. J. Biosci. 2002, 27, 703–714. [Google Scholar] [CrossRef]
- Byung-Sung, P. Effect of oral administration of Jerusalem artichoke inulin on reducing blood lipid and glucose in STZ-Induced diabetic rats. J. Anim.Vet. Adv. 2011, 10, 2501–2507. [Google Scholar] [CrossRef]
- Sawicka, B.; Skiba, D.; Pszczółkowski, P.; Aslan, I.; Sharifi-Rad, J.; Krochmal-Marczak, B. Jerusalem artichoke (H. tuberosus L.) as a medicinal plant and its natural products. Cell. Mol. Biol. 2020, 66, 160–177. [Google Scholar] [CrossRef]
- Nair, K.K.; Kharb, K.; Thompkinson, D.K. Inulin dietary fiber with functional and health attributes. Food Rev. Int. 2010, 26, 189–203. [Google Scholar] [CrossRef]
- Kim, H.S.; Han, G.D. Hypoglycemic and hepatoprotective effects of Jerusalem artichoke extracts on streptozotocin-induced diabetic rats. Food Sci. Biotechnol. 2013, 22, 1121–1124. [Google Scholar] [CrossRef]
- Pisonero-Vaquero, S.; González-Gallego, J.; Sánchez-Campos, S.; García-Mediavilla, M.V. Flavonoids and related compounds in Non-Alcoholic Fatty Liver Disease therapy. Curr. Med. Chem. 2015, 22, 2991–3012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Cai, G.; Sun, X.; Chen, X. Treatment of chronic kidney disease using a traditional Chinese medicine, Flos Abelmoschus manihot (Linnaeus) Medicus (Malvaceae). Clin. Exp. Pharm. Physiol. 2016, 43, 145–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eugenio-Pérez, D.; Medina-Fernandez, L.Y.; Saldivar-Anaya, J.A.; Molina-Jijon, E.; Pedraza-Chaverri, J. Role of dietary antioxidant agents in chronic kidney disease. Free Radic. Dis. 2016, 1, 331–350. [Google Scholar]
- Rodriguez-Ramiro, I.; Vauzour, D.; Minihane, A.M. Polyphenols and non-alcoholic fatty liver disease: Impact and mechanisms. Proc. Nutr. Soc. 2016, 75, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T. Catechin treatment ameliorates diabetes and its complications in streptozotocin-induced diabetic rats. Dose-Response 2017, 15, 1559325817691158. [Google Scholar] [CrossRef]
- Li, H.; Yao, Y.; Li, L. Coumarins as potential antidiabetic agents. J. Pharm. Pharmacol. 2017, 69, 1253–1264. [Google Scholar] [CrossRef] [Green Version]
- Vargas, F.; Romecín, P.; García-Guillén, A.I.; Wangesteen, R.; Vargas-Tendero, P.; Paredes, M.D.; Atucha, N.M.; García-Estañ, J. Flavonoids in kidney health and disease. Front. Physiol. 2018, 24, 9–394. [Google Scholar] [CrossRef] [Green Version]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Liakopoulos, V.; Roumeliotis, S.; Bozikas, A.; Eleftheriadis, T.; Dounousi, E. Antioxidant supplementation in renal replacement therapy patients: Is there evidence? Oxid. Med. Cell. Longev. 2019, 2019, 9109473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, N.; Chen, F.; Dai, D. The regulation of host intestinal microbiota by polyphenols in the development and prevention of chronic kidney disease. Front. Immunol. 2020, 10, 2981–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, A.K.; Mulpuru, V.; Mishra, N. Discovery of novel coumarin analogs against the α-glucosidase protein target of Diabetes mellitus: Pharmacophore-based QSAR, docking, and molecular dynamics simulation studies. ACS Omega 2020, 5, 32234–32249. [Google Scholar] [CrossRef] [PubMed]
- Ritsema, T.; Smeekens, S.C. Engineering fructan metabolism in plants. J. Plant Physiol. 2003, 160, 811–820. [Google Scholar] [CrossRef]
- Abdel Raoof, G.F.; Mohamed, K.Y. Natural Products for the Management of Diabetes. In Studies in Natural Products Chemistry; Atta-ur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 59, pp. 323–374. [Google Scholar]
- Xu, H.; Liang, M.; Xu, L.; Li, H.; Zhang, X.; Kang, J.; Zhao, Q.; Zhao, H. Cloning and functional characterization of two abiotic stress-responsive Jerusalem artichoke (Helianthus tuberosus) fructan 1-exohydrolases (1-FEHs). Plant Mol. Biol. 2015, 87, 81–98. [Google Scholar] [CrossRef]
- Van den Ende, W. Novel fructan exohydrolase: Unique properties and applications for human health. J. Exp. Bot. 2018, 69, 4227–4231. [Google Scholar] [CrossRef]
- Saengkanuk, A.; Nuchadomrong, S.; Jogloy, S.; Patanothai, A.; Srijaranai, S. A simplified spectrophotometric method for the determination of inulin in Jerusalem artichoke (Helianthus tuberosus L.) tubers. Eur. Food Res. Technol. 2011, 233, 609–616. [Google Scholar] [CrossRef]
- Ahn, C.S.; Lee, J.H.; Reum Hwang, A.; Kim, W.T.; Pai, H.S. Prohibitin is involved in mitochondrial biogenesis in plants. Plant J. 2006, 46, 658–667. [Google Scholar] [CrossRef]
- Belser, M.; Walker, D.W. Role of prohibitins in aging and therapeutic potential against age-related diseases. Front. Genet. 2021, 12, 714228. [Google Scholar] [CrossRef]
- Dong, W.Q.; Chao, M.; Lu, Q.H.; Chai, W.L.; Zhang, W.; Chen, X.Y.; Liang, E.S.; Wang, L.B.; Tian, H.L.; Chen, Y.G.; et al. Prohibitin overexpression improves myocardial function in diabetic cardiomyopathy. Oncotarget 2016, 7, 66–80. [Google Scholar] [CrossRef] [Green Version]
- Kakarla, M.; Puppala, V.K.; Tyagi, S.; Anger, A.; Repp, K.; Wang, J.; Ying, R.; Widlansky, M.E. Circulating levels of mitochondrial uncoupling protein 2, but not prohibitin, are lower in humans with type 2 diabetes and correlate with brachial artery flow-mediated dilation. Cardiovasc. Diabetol. 2019, 18, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Koushyar, S.; Jiang, W.G.; Dart, D.A. Unveiling the potential of prohibitin in cancer. Cancer Lett. 2015, 369, 316–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, P.E. Peptidyl-prolyl isomerases: A new twist to transcription. EMBO Rep. 2002, 3, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pliyev, B.K.; Gurvits, B.Y. Peptidyl-prolyl cis-trans isomerases: Structure and functions. Biochem. Biokhimiia 1999, 64, 738–751. [Google Scholar]
- Ingelsson, B.; Shapiguzov, A.; Kieselbach, T.; Vener, A.V. Peptidyl-prolyl isomerase activity in chloroplast thylakoid lumen is a dispensable function of immunophilins in Arabidopsis thaliana. Plant Cell Physiol. 2009, 50, 1801–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreiber, S.L. Chemistry and biology of the immunophilins and their immunosuppressive ligands. Science 1991, 251, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, M.; Kosaka, K.; Tsushima, S.; Uchida, C.; Takahashi, K.; Takahashi, N.; Tsubuki, M.; Hara, Y.; Uchida, T. Food polyphenols targeting peptidyl prolyl cis/trans isomerase Pin1. Biochem. Biophys. Res. Commun. 2018, 499, 681–687. [Google Scholar] [CrossRef]
- Durst, F. The correlation of phenylalanine ammonia-lyase and cinnamic acid-hydroxylase activity changes in Jerusalem artichoke tuber tissues. Planta 1976, 132, 221–227. [Google Scholar] [CrossRef]
- Şirin, S.; Aydaş, S.B.; Aslım, B. Biochemical evaluation of phenylalanine ammonia lyase from endemic plant Cyathobasis fruticulosa (Bunge) Aellen. for the dietary treatment of phenylketonuria. Food Technol. Biotechnol. 2016, 54, 296–303. [Google Scholar] [CrossRef]
- Eissa, H.A.; Abdallah, Z.Y.; Khalil, W.K.B.; Ibrahim, W.A.; Booles, H.F.; Hassanane, M.M. Effect of natural PAL-enzyme on the quality of egg white and mushroom flour and study its impact on the expression of PKU related genes and phenylalanine reduction in mice fed on. J. Genet. Eng. Biotechnol. 2017, 15, 443–451. [Google Scholar] [CrossRef]
- Bourne, Y.; Zamboni, V.; Barre, A.; Peumans, W.J.; Van Damme, E.J.; Rougé, P. Helianthus tuberosus lectin reveals a widespread scaffold for mannose-binding lectins. Structure 1999, 7, 1473–1482. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Li, Y.; Xue, C.; Dong, N.; Bi, C.; Shan, A. Aquaporin: Targets for dietary nutrients to regulate intestinal health. J. Anim. Physiol. Anim. Nutr. 2022, 106, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.; Mejia, S.; Vojdani, A. Plant and human aquaporins: Pathogenesis from gut to brain. Immunol. Res. 2019, 67, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauh, G.Y.; Fischer, A.M.; Grimes, H.D.; Ryan, C.A.; Rogers, J.C., Jr. Delta-Tonoplast intrinsic protein defines unique plant vacuole functions. Proc. Natl. Acad. Sci. USA 1998, 95, 12995–12999. [Google Scholar] [CrossRef] [Green Version]
- Chardin, C.; Schenk, S.T.; Hirt, H.; Colcombet, J.; Krapp, A. Review: Mitogen-activated protein kinases in nutritional signaling in Arabidopsis. Plant Sci. 2017, 260, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Daldin, M.; Fodale, V.; Cariulo, C.; Azzollini, L.; Verani, M.; Martufi, P.; Spiezia, M.C.; Deguire, S.M.; Cherubini, M.; Macdonald, D.; et al. Polyglutamine expansion affects huntingtin conformation in multiple Huntington’s disease models. Sci. Rep. 2017, 7, 5070. [Google Scholar] [CrossRef] [Green Version]
- Waelter, S.; Scherzinger, E.; Hasenbank, R.; Nordhoff, E.; Lurz, R.; Goehler, H.; Gauss, C.; Sathasivam, K.; Bates, G.P.; Lehrach, H.; et al. The huntingtin interacting protein HIP1 is a clathrin and alpha-adaptin-binding protein involved in receptor-mediated endocytosis. Hum. Mol. Genet. 2001, 10, 1807–1817. [Google Scholar] [CrossRef] [Green Version]
- Imai, Y.; Soda, M.; Hatakeyama, S.; Akagi, T.; Hashikawa, T.; Nakayama, K.I.; Takahashi, R. CHIP is associated with Parkin, a gene responsible for familial Parkinson’s disease, and enhances its ubiquitin ligase activity. Mol. Cell 2002, 10, 55–67. [Google Scholar] [CrossRef]
- Kurepa, J.; Wang, S.; Li, Y.; Smalle, J. Proteasome regulation, plant growth and stress tolerance. Plant Signal. Behav. 2009, 4, 924–927. [Google Scholar] [CrossRef] [Green Version]
- Valdés-Ramos, R.; Guadarrama-López, A.L.; Martínez-Carrillo, B.E.; Benítez-Arciniega, A.D. Vitamins and type 2 diabetes mellitus. Endocr. Metab. Immune Disord. Drug Targets 2015, 15, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Smolek, M.K.; Notaroberto, N.F.; Jaramillo, A.G.; Pradillo, L.R. Intervention with vitamins in patients with nonproliferative diabetic retinopathy: A pilot study. Clin. Ophthalmol. 2013, 7, 1451–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unoki-Kubota, H.; Yamagishi, S.; Takeuchi, M.; Bujo, H.; Saito, Y. Pyridoxamine, an inhibitor of advanced glycation end product (AGE) formation ameliorates insulin resistance in obese, type 2 diabetic mice. Protein Pept. Lett. 2010, 17, 1177–1181. [Google Scholar] [CrossRef] [PubMed]
- Al-Attas, O.S.; Al-Daghri, N.M.; Alfadda, A.A.; Abd-Alrahman, S.H.; Sabico, S. Blood thiamine and its phosphate esters as measured by high-performance liquid chromatography: Levels and associations in diabetes mellitus patients with varying degrees of microalbuminuria. J. Endocrinol. Investig. 2012, 35, 951–956. [Google Scholar]
- Gonzalez-Ortiz, M.; Martinez-Abundis, E.; Robles-Cervantes, J.A.; Ramirez-Ramirez, V.; Ramos-Zavala, M.G. Effect of thiamine administration on metabolic profile, cytokines and inflammatory markers in drug-naive patients with type 2 diabetes. Eur. J. Nutr. 2010, 50, 145–149. [Google Scholar] [CrossRef]
- Pittas, A.G.; Lau, J.; Hu, F.B.; Dawson-Hughes, B. The role of vitamin D and calcium in type 2 diabetes. A systematic review and meta-analysis. J. Clin. Endocrinol. Metab. 2007, 92, 2017–2029. [Google Scholar] [CrossRef] [PubMed]
- Bellan, M.; Guzzaloni, G.; Rinaldi, M.; Merlotti, E.; Ferrari, C.; Tagliaferri, A.; Pirisi, M.; Aimaretti, G.; Scacchi, M.; Marzullo, P. Altered glucose metabolism rather than naive type 2 diabetes mellitus (T2DM) is related to vitamin D status in severe obesity. Cardiovasc. Diabetol. 2014, 13, 57–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wu, L.; Du, S.; Hu, Y.; Fan, Y.; Ma, J. 1,25(OH)2D3 inhibits high glucose-induced apoptosis and ROS production in human peritoneal mesothelial cells via the MAPK/P38 pathway. Mol. Med. Rep. 2016, 14, 839–844. [Google Scholar] [CrossRef] [Green Version]
- Arai, C.; Arai, N.; Mizote, A.; Kohno, K.; Iwaki, K.; Hanaya, T.; Arai, S.; Ushio, S.; Fukuda, S. Trehalose prevents adipocyte hypertrophy and mitigates insulin resistance. Nutr. Res. 2010, 30, 840–848. [Google Scholar] [CrossRef]
- Pan, Y.; Carroll, J.D.; Asano, N.; Pastuszak, I.; Edavana, V.K.; Elbein, A.D. Trehalose synthase converts glycogen to trehalose. FEBS J. 2008, 275, 3408–3420. [Google Scholar] [CrossRef]
- Joseph, J.A.; Akkermans, S.; Nimmegeers, P.; Van Impe, J.F.M. Bioproduction of the recombinant sweet protein thaumatin: Current State of the Art and Perspectives. Front. Microbiol. 2019, 10, 695–713. [Google Scholar] [CrossRef] [Green Version]
- Geraldes, P.; King, G.L. Activation of protein kinase C isoforms and its impact on diabetic complications. Circ. Res. 2010, 106, 1319–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menne, J.; Meier, M.; Park, J.; Haller, H. Inhibition of protein kinase C in diabetic nephropathy—Where do we stand? Nephrol. Dial. Transplant. 2009, 24, 2021–2023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skvortsova, I. Profilin 1: Do we have a novel proteomefound biomarker predicting response to anticancer therapy? Proteomics 2013, 13, 2069–2071. [Google Scholar] [CrossRef] [PubMed]
- Pimm, M.L.; Hotaling, J.; Henty-Ridilla, J.L. Profilin choreographs actin and microtubules in cells and cancer. Int. Rev. Cell Mol. Biol. 2020, 355, 155–204. [Google Scholar]
- Dráber, P.; Dráberová, E. Dysregulation of Microtubule Nucleating Proteins in Cancer Cells. Cancers 2021, 13, 5638. [Google Scholar] [CrossRef]
- Yao, W.; Ji, S.; Qin, Y.; Yang, J.; Xu, J.; Zhang, B.; Xu, W.; Liu, J.; Shi, S.; Liu, L.; et al. Profilin-1 suppresses tumorigenicity in pancreatic cancer through regulation of the SIRT3-HIF1α axis. Mol. Cancer 2014, 13, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Styskal, J.; Remmen, V.; Richardson, H.A.; Salmon, A.B. Oxidative stress and diabetes: What can we learn about insulin resistance from antioxidant mutant mouse models? Free Radic. Biol. Med. 2012, 52, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Wahab, N.A.; Abdullah, N.; Aminudin, N. Characterisation of potential antidiabetic-related proteins from Pleurotus pulmonarius (Fr.) Quél. (Grey oyster mushroom) by MALDI-TOF/TOF mass spectrometry. BioMed Res. Int. 2014, 2014, 131607. [Google Scholar] [CrossRef] [Green Version]
- Beisswenger, P.J.; Howell, S.K.; Smith, K.; Szwergold, B.S. Glyceraldehyde-3-phosphate dehydrogenase activity as an independent modifier of methylglyoxal levels in diabetes. Biochim. Biophys. Acta Mol. Basis Dis. 2003, 1637, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Maher, P. Methylglyoxal, advanced glycation end products and autism: Is there a connection? Med. Hypotheses 2012, 78, 548–552. [Google Scholar] [CrossRef]
- Malinova, I.; Kunz, H.H.; Alseekh, S.; Herbst, K.; Fernie, A.R.; Gierth, M.; Fettke, J. Reduction of the cytosolic phosphoglucomutase in Arabidopsis reveals impact on plant growth, seed and root development, and carbohydrate partitioning. PLoS ONE 2014, 9, e112468. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.; Marshall, J.; Milne, D.; Kanhai, J.; Kantikar, S.M.; Raju, S. Hypoglycaemic activity of Chrysobalanus icaco (fat-pork) fruit extract in diabetes induced rats, Asian. J. Pharm. Biol. Res. 2011, 1, 512–517. [Google Scholar]
- Kobayashi, H.; Suzuki, M.; Kanayama, N.; Terao, T. A soybean Kunitz trypsin inhibitor suppresses ovarian cancer cell invasion by blocking urokinase up regulation. Clin. Exp. Metastasis 2004, 21, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, S.; Chen, Z. Plant protease inhibitors in therapeutics-focus on cancer therapy. Front. Pharmacol. 2016, 7, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leng, F.; Wu, L.Y.; Lu, C.; Pan, X.M. Determinants of thermostability in serine hydroxy methyl transferase identified by principal component analysis. Sci. Rep. 2017, 7, 46463. [Google Scholar] [CrossRef] [Green Version]
- Kulak, N.A.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, encapsulated proteomic-sample processing applied to copy-number estimation in eukaryotic cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef]
- Pajarillo, E.A.; Kim, S.H.; Lee, J.Y.; Valeriano, V.D.; Kang, D.K. Quantitative proteogenomics and the reconstruction of the metabolic pathway in Lactobacillus mucosae LM1. Korean J. Food Sci. Anim. Resour. 2015, 35, 692–702. [Google Scholar] [CrossRef] [Green Version]
- Min, C.W.; Lee, S.H.; Cheon, Y.E.; Han, W.Y.; Ko, J.M.; Kang, H.W.; Kim, Y.C.; Agrawal, G.K.; Rakwal, R.; Gupta, R.; et al. In-depth proteomic analysis of Glycine max seeds during controlled deterioration treatment reveals a shift in seed metabolism. J. Proteom. 2017, 169, 125–135. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Deutsch, E.W.; Bandeira, N.; Sharma, V.; Perez-Riverol, Y.; Carver, J.J.; Kundu, D.J.; García-Seisdedos, D.; Jarnuczak, A.F.; Hewapathirana, S.; Pullman, B.S.; et al. The ProteomeXchange consortium in 2020: Enabling ‘big data’ approaches in proteomics. Nucleic Acids Res. 2020, 48, D1145–D1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Riverol, Y.; Xu, Q.W.; Wang, R.; Szkoreit, J.; Griss, J.; Sanchez, A.; Reisinger, F.; Ternent, T.; del-Toro, N.; Dianes, J.A.; et al. PRIDE Inspector Toolsuite: Moving towards a universal visualization tool for proteomics data standard formats and quality assessment of ProteomeXchange datasets. Mol. Cell. Proteom. 2016, 15, 305–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]






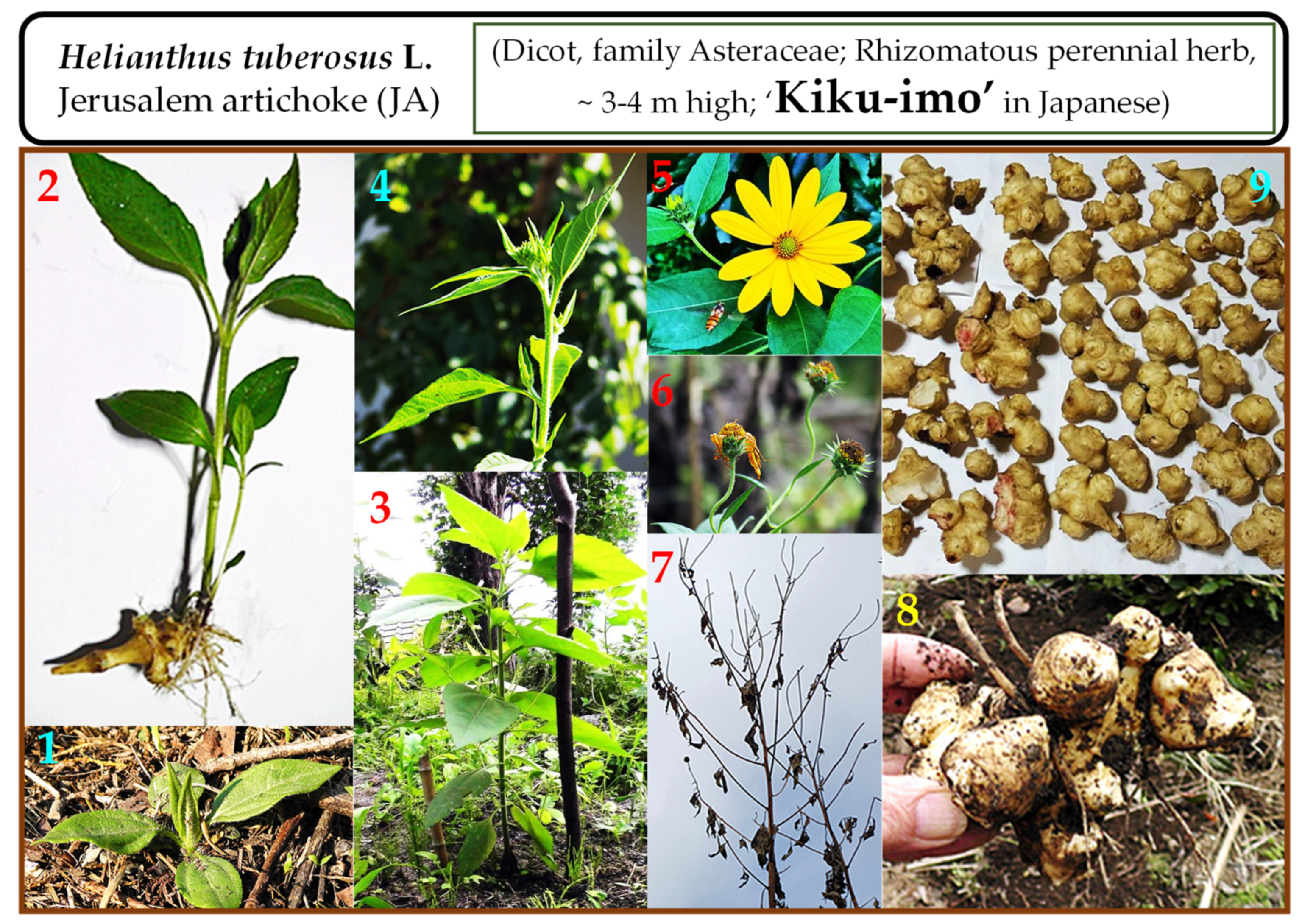{kind=link}
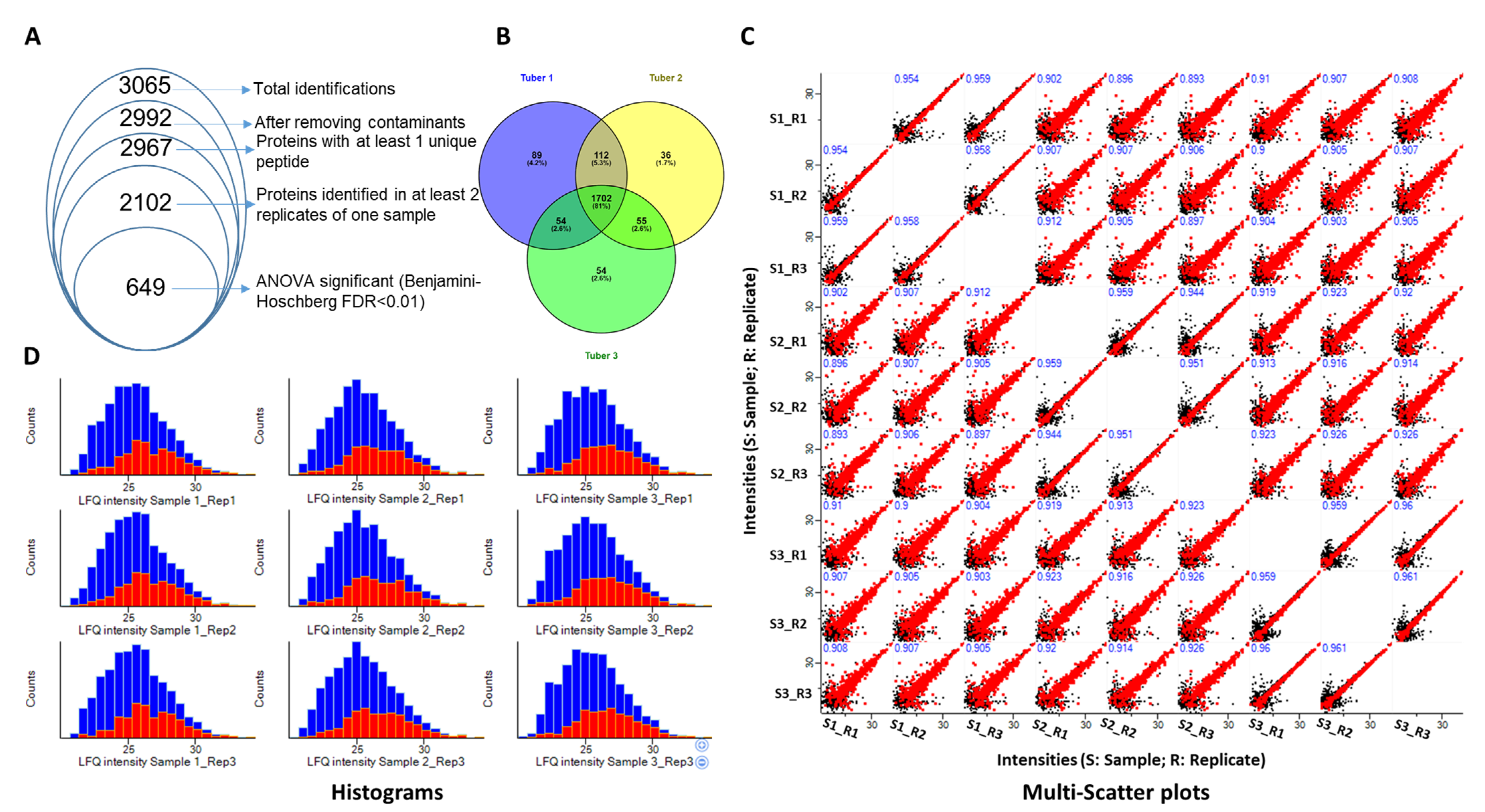{kind=link}
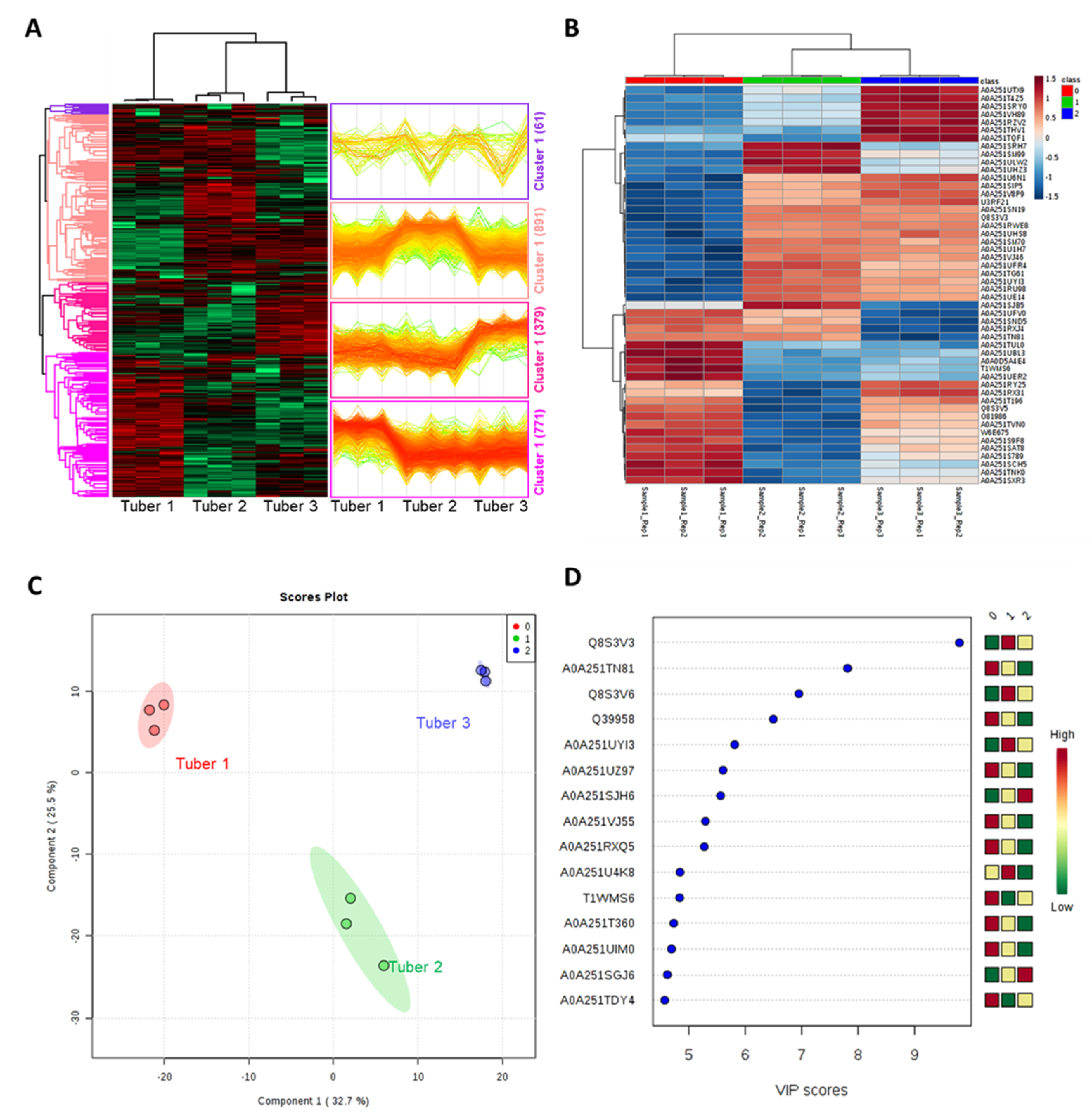{kind=link}
{kind=link}
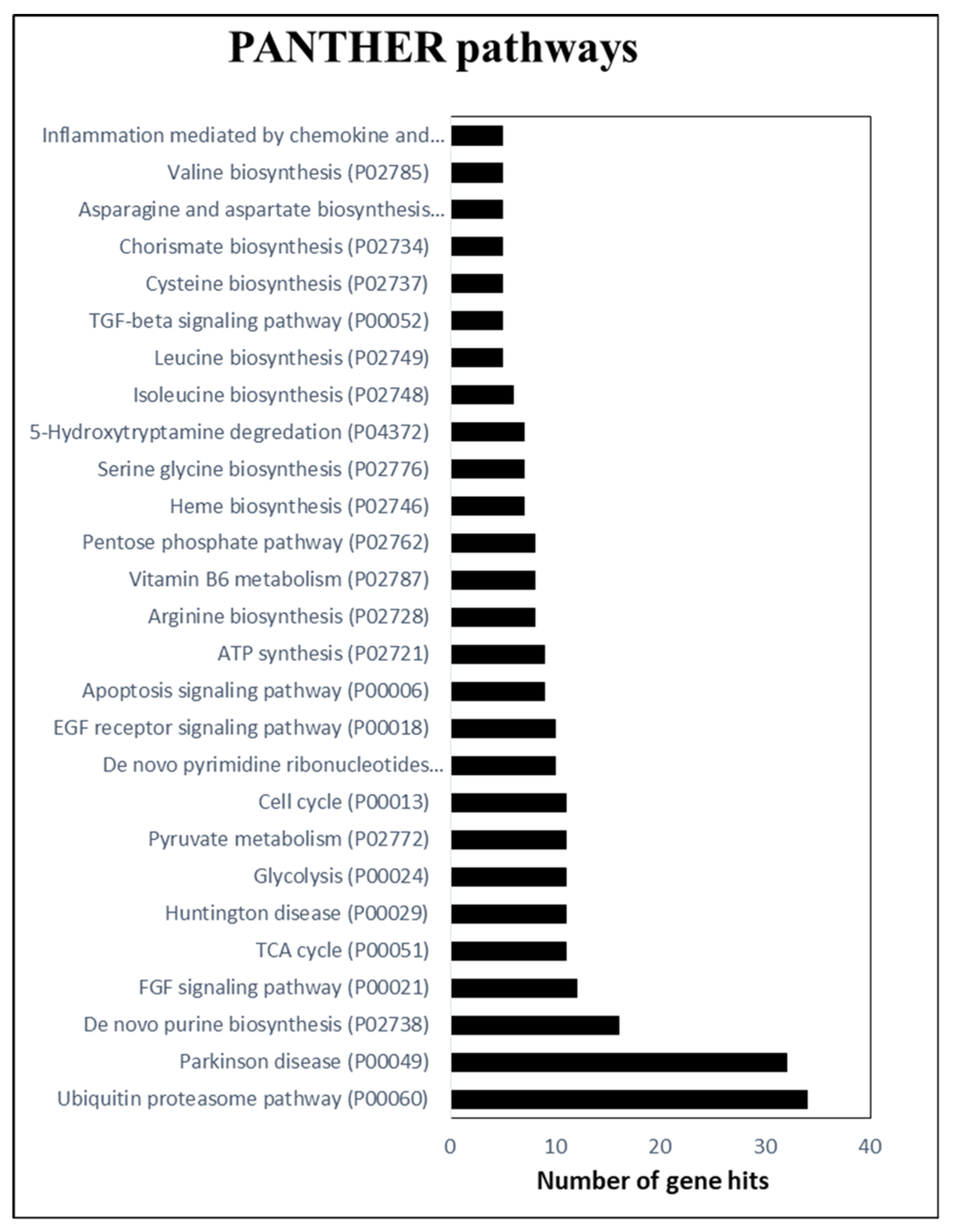{kind=link}
| Sample | Rep_1 | Rep_2 | Rep_3 | Common Proteins in All Replicates | Standard Deviation | Replication Ratio | Percentage Coefficient of Variation (CV%) |
|---|---|---|---|---|---|---|---|
| Sample 1 | 1917 | 1966 | 1955 | 1780 | 25.71 | 0.915 | 8.5 |
| Sample 2 | 1901 | 1898 | 1920 | 1732 | 11.93 | 0.909 | 9.1 |
| Sample 3 | 1887 | 1917 | 2439 | 1701 | 310.4 | 0.817 | 18.26 |
| S.No | Protein IDs | Annotation | No of Peptides |
|---|---|---|---|
| 1 | A0A251UTX9 | Phospho-2-dehydro-3-deoxyheptonate aldolase | 12 |
| 2 | A0A251T4Z5 | Putative caffeoyl-CoA O-methyltransferase | 8 |
| 3 | A0A251SRY0 | Phenylalanine ammonia-lyase | 16 |
| 4 | A0A251VH89 | Phenylalanine ammonia-lyase | 17 |
| 5 | A0A251RZV2 | Putative bifunctional polymyxin resistance protein, ArnA | 6 |
| 6 | A0A251THV1 | Phospho-2-dehydro-3-deoxyheptonate aldolase | 13 |
| 7 | A0A251TQF1 | Putative FAD-binding Berberine family protein; Belongs to the oxygen-dependent FAD-linked oxidoreductase family. | 6 |
| 8 | A0A251SRH7 | Putative cobalamin-independent methionine synthase | 45 |
| 9 | A0A251SM99 | Putative heat shock protein Hsp90 family | 31 |
| 10 | A0A251ULW2 | Putative glucose/ribitol dehydrogenase; Belongs to the short-chain dehydrogenases/reductases (SDR) family. | 6 |
| 11 | A0A251UHZ3 | Glutathione peroxidase; Belongs to the glutathione peroxidase family. | 5 |
| 12 | A0A251U6N1 | Putative caffeic acid 3-O-methyltransferase; Belongs to the class I-like SAM-binding methyltransferase superfamily. Cation-independent O-methyltransferase family. | 9 |
| 13 | A0A251SIP5 | UDP-glucose 6-dehydrogenase | 13 |
| 14 | A0A251V8P9 | Putative triose phosphate/phosphoenolpyruvate translocator | 2 |
| 15 | U3RF21 | S-adenosylmethionine synthase | 16 |
| 16 | A0A251SN19 | Peroxidase; Removal of H(2)O(2), oxidation of toxic reductants, biosynthesis and degradation of lignin, suberization, auxin catabolism, response to environmental stresses such as wounding, pathogen attack and oxidative stress. | 7 |
| 17 | Q8S3V3 | Tuber agglutinin | 12 |
| 18 | A0A251RWE8 | Putative heat shock protein; Belongs to the ClpA/ClpB family. | 35 |
| 19 | A0A251UHS8 | Methylenetetrahydrofolate reductase; Belongs to the methylenetetrahydrofolate reductase family. | 12 |
| 20 | A0A251SM70 | Putative pyridoxine biosynthesis 1.2; Belongs to the PdxS/SNZ family. | 6 |
| 21 | A0A251U1H7 | Putative HSP20-like chaperone; Belongs to the small heat shock protein (HSP20) family. | 4 |
| 22 | A0A251VJ46 | S-adenosylmethionine synthase; Catalyzes the formation of S-adenosylmethionine from methionine and ATP. | 12 |
| 23 | A0A251UFR4 | Putative 22.0 kDa class IV heat shock protein; Belongs to the small heat shock protein (HSP20) family. | 7 |
| 24 | A0A251TG61 | Peroxidase; Removal of H(2)O(2), oxidation of toxic reductants, biosynthesis and degradation of lignin, suberization, auxin catabolism, response to environmental stresses such as wounding, pathogen attack and oxidative stress. | 10 |
| 25 | A0A251UYI3 | Peptidylprolyl isomerase | 9 |
| 26 | A0A251RU98 | Putative HSP20-like chaperone; Belongs to the small heat shock protein (HSP20) family. | 7 |
| 27 | A0A251UE14 | Putative casein lytic proteinase B3; Belongs to the ClpA/ClpB family. | 23 |
| 28 | A0A251SJB5 | Putative sieve element occlusion | 10 |
| 29 | A0A251UFV0 | Putative P-loop containing nucleoside triphosphate hydrolases superfamily protein | 16 |
| 30 | A0A251SND5 | Putative RNA-binding (RRM/RBD/RNP motifs) family protein | 6 |
| 31 | A0A251RXJ4 | SUMO-activating enzyme subunit; Belongs to the ubiquitin-activating E1 family. | 4 |
| 32 | A0A251TN81 | Peptidyl-prolyl cis-trans isomerase; PPIases accelerate the folding of proteins. It catalyzes the cis-trans isomerization of proline imidic peptide bonds in oligopeptides; Belongs to the cyclophilin-type PPIase family. | 6 |
| 33 | A0A251TUL0 | Putative peptidase C13, legumain | 4 |
| 34 | A0A251UBL3 | Putative START-like domain-containing protein | 8 |
| 35 | A0A0D5A4E4 | Fructan 1-exohydrolase | 6 |
| 36 | T1WMS6 | GLYCENE RICH RNA BINING PROTEIN | 4 |
| 37 | A0A251UER2 | Putative alpha-amylase/subtilisin inhibitor | 5 |
| 38 | A0A251RY25 | Putative granulin; Belongs to the peptidase C1 family. | 4 |
| 39 | A0A251RX31 | Putative aldehyde dehydrogenase 2B4; Belongs to the aldehyde dehydrogenase family. | 19 |
| 40 | A0A251T196 | Putative cupredoxin | 4 |
| 41 | Q8S3V5 | TUBER AGGULTIN | 8 |
| 42 | O81986 | SUCROSE FRUCTOSYL TRASFERASE | 12 |
| 43 | A0A251S9F8 | Putative eukaryotic aspartyl protease family protein; Belongs to the peptidase A1 family. | 4 |
| 44 | A0A251SAT8 | UTP--glucose-1-phosphate uridylyltransferase | 22 |
| 45 | A0A251S789 | Prohibitin | 11 |
| 46 | A0A251SCH5 | Peptidylprolyl isomerase | 3 |
| 47 | A0A251TNK0 | Putative cytochrome P450; Belongs to the cytochrome P450 family. | 6 |
| 48 | A0A251SXR3 | Putative oxysterol-binding family protein; Belongs to the OSBP family. | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakku, R.K.; Gupta, R.; Min, C.-W.; Kim, S.-T.; Takahashi, G.; Shibato, J.; Shioda, S.; Takenoya, F.; Agrawal, G.K.; Rakwal, R. Unravelling the Helianthus tuberosus L. (Jerusalem Artichoke, Kiku-Imo) Tuber Proteome by Label-Free Quantitative Proteomics. Molecules 2022, 27, 1111. https://doi.org/10.3390/molecules27031111
Bakku RK, Gupta R, Min C-W, Kim S-T, Takahashi G, Shibato J, Shioda S, Takenoya F, Agrawal GK, Rakwal R. Unravelling the Helianthus tuberosus L. (Jerusalem Artichoke, Kiku-Imo) Tuber Proteome by Label-Free Quantitative Proteomics. Molecules. 2022; 27(3):1111. https://doi.org/10.3390/molecules27031111
Chicago/Turabian StyleBakku, Ranjith Kumar, Ravi Gupta, Cheol-Woo Min, Sun-Tae Kim, Genboku Takahashi, Junko Shibato, Seiji Shioda, Fumiko Takenoya, Ganesh Kumar Agrawal, and Randeep Rakwal. 2022. "Unravelling the Helianthus tuberosus L. (Jerusalem Artichoke, Kiku-Imo) Tuber Proteome by Label-Free Quantitative Proteomics" Molecules 27, no. 3: 1111. https://doi.org/10.3390/molecules27031111